
01 Scientia 9 2 - Inicio | Centro Universitario de Ciencias ... Luz Elena Claudio G., Patricia...
76
Efecto de la aplicación de agroquímicos y abonos orgánicos sobre la producción de frijol y la población microbiana del suelo Manuel Morales Torres, Rogelio Lépiz Ildefonso, Diego R. González Eguiarte, Ricardo Nuño Romero, Luz Elena Claudio G., Patricia Zarazúa Villaseñor, Salvador Mena Munguía y Eduardo López Alcocer 81 Sistemas silvopastoriles para una reconversión ambiental de la ganadería en la cuenca del río San Pedro en Camagüey, Cuba Zoe G. Acosta Gutiérrez 95 Extracción de ADN en accesiones de maíz y teocintle Martha Isabel Torres-Morán, Moisés Martín Morales-Rivera, José de Jesús Sánchez-González, Lino De la Cruz-Larios, José Ron-Parra y Fernando Santacruz-Ruvalcaba 105 Selección por comportamiento per se en líneas de jitomate: producción de fruto y semilla Eduardo Rodríguez Guzmán, Aquiles Carballo Carballo, Gustavo Baca Castillo, Rogelio Lépiz Ildefonso, Lino de la Cruz Larios y Salvador Mena Munguía 119 El factor gametofítico-1 (ga1) en teocintle (Zea spp.) en México Lino De la Cruz Larios, José de Jesús Sánchez González, José Guadalupe Rodríguez Flores, José Ron Parra y Fernando Santacruz-Ruvalcaba 129 CONTENIDO ISSN 1665-8493 CUCBA scientia Universidad de Guadalajara COORDINACIÓN DE I NVESTIGACIÓN DICIEMBRE DE 2007 VOLUMEN NÚMERO 9 2 Centro Universitario de Ciencias Biológicas y Agropecuarias AHORA ¡ en línea ! continúa en la contraportada R E V I S T A C I E N T Í F I C A FECHA EFECTIVA DE PUBLICACIÓN 29 DE DICIEMBRE DE 2007
Transcript of 01 Scientia 9 2 - Inicio | Centro Universitario de Ciencias ... Luz Elena Claudio G., Patricia...

Efecto de la aplicación de agroquímicos y abonos orgánicos sobre la producción de frijol y la población microbiana del sueloManuel Morales Torres, Rogelio Lépiz Ildefonso, Diego R. González Eguiarte, Ricardo Nuño Romero, Luz Elena Claudio G., Patricia Zarazúa Villaseñor, Salvador Mena Munguía y Eduardo López Alcocer 81
Sistemas silvopastoriles para una reconversión ambiental de la ganadería en la cuenca del río San Pedro en Camagüey, CubaZoe G. Acosta Gutiérrez 95
Extracción de ADN en accesiones de maíz y teocintleMartha Isabel Torres-Morán, Moisés Martín Morales-Rivera, José de Jesús Sánchez-González, Lino De la Cruz-Larios, José Ron-Parra y Fernando Santacruz-Ruvalcaba 105
Selección por comportamiento per se en líneas de jitomate: producción de fruto y semillaEduardo Rodríguez Guzmán, Aquiles Carballo Carballo, Gustavo Baca Castillo, Rogelio Lépiz Ildefonso, Lino de la Cruz Larios y Salvador Mena Munguía 119
El factor gametofítico-1 (ga1) en teocintle (Zea spp.) en MéxicoLino De la Cruz Larios, José de Jesús Sánchez González, José Guadalupe Rodríguez Flores, José Ron Parra y Fernando Santacruz-Ruvalcaba 129
CONTENIDO
ISSN 1665-8493
CUCBAscientiaUniversidad de
Guadalajara
COORDINACIÓN DEINVESTIGACIÓN
DICI
EMBR
E DE
200
7VOLUMEN
NÚMERO
92
Centro Universitario deCiencias Biológicas y
Agropecuarias
AHORA
¡en lín
ea!
continúa en la contraportada
R E V I S T A C I E N T Í F I C A
FECHA EFECTIVA DE PUBLICACIÓN 29 DE DICIEMBRE DE 2007

DICI
EMBR
E DE
200
7VOLUMEN
NÚMERO
92
Efecto de la aplicación de compostas sobre las características físicas, químicas y biológicas del suelo en el cultivo de frijolMartha Delia Ocegueda Reyes, Eduardo López Alcocer, Rogelio Lépiz Ildefonso, Diego R. González Eguiarte, Ricardo Nuño Romero, Fernando López Alcocer y Patricia Zarazúa Villaseñor 143
CONTENIDO (continuación)
Consulte esta y otras publicaciones vía internet enwww.cucba.udg.mx/new/publicaciones
U N I V E R S I D A D D E G U A D A L A J A R A
CENTRO UNIVERSITARIO DE CIENCIAS BIOLÓGICAS Y AGROPECUARIAS

scientia-CUCBA es el órgano ofi cial de difusión científi ca del Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA) de la Universidad de Guadalajara. Es una publicación interdisciplinaria de ciencias biológicas, agropecuarias y ambientales que se da a la luz cada seis meses.
Editor Jefe: Servando Carvajal.
Comité Editorial
Óscar Aguirre Calderón. Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León, México. [email protected]
Esther Albarrán Rodríguez. Departamento de Medicina Veterinaria, CUCBA. Zapopan, México.
[email protected] Bañuelos Pineda. Departamento de Medicina
Veterinaria. CUCBA. Zapopan, Jalisco, México. [email protected]
Anatoli Borodanienko. Instituto de Ciencias Agrícolas (ICA), Universidad de Guanajuato, México.
[email protected] Caballero Chacón. Departamento de Fisiología
y Farmacología, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México, México. [email protected]
Miguel Ángel Carmona Medero. Facultad de Veterinaria. Universidad Nacional Autónoma de México. México. [email protected]
Servando Carvajal. Departamento de Botánica y Zoología CUCBA. Zapopan, Jalisco, México.
[email protected] Castillo Ayala. Universidad de Texas A & M.
U.S.A. [email protected] Eduardo Chalita Tovar. Colegio de Posgraduados,
Chapingo, México. [email protected]. Daniel G. Debouck. Centro Internacional de Agricultura
Tropical (CIAT), Cali, Colombia. [email protected] Esteve. Centro Mas Bove. Tarragona, España. [email protected]é Funes Rodríguez. CICIMAR-IPN, Baja California,
México. [email protected] García Moya. Colegio de Posgraduados de
Chapingo, México. [email protected] Gliesman. Universidad de California, Campus Santa
Cruz. usa. [email protected] Gómez Kasky. Universidad de Las Villas, Cuba.
[email protected] González Izquierdo. Universidad de Pinar del Río,
Cuba. [email protected] Hansen. Midamerican International Agricultural
Consortium, Iowa State University. U.S.A. [email protected] Lemus Flores. Instituto de Medicina Veterinaria
de la Universidad Autónoma de Nayarit. México. [email protected]
Rogelio Lépiz Ildefonso. Departamento de Producción Agrícola, CUCBA. Zapopan, Jalisco, México. [email protected]
Ángel Luque Escalona. Universidad de Las Palmas, Gran Canaria, España. [email protected]
Jorge Manzo. Laboratorio de Neurociencias, Universidad Veracruzana, México. [email protected]
Jaime Morales Hernández. Programa Universitario de Medio Ambiente y Desarrollo. Instituto Tecnológico de Estudios Superiores de Occidente (ITESO), Jalisco, México. [email protected]
Xavier Navarro Acebes. Departament de Biología Cel·lular, de Fisiología i d’immunologia, Bellaterra, España.
[email protected] A. Navarro. Centro Agronómico Tropical de
Investigación y Enseñanza (CATIE) - Turrialba, Costa Rica. [email protected]
Michael F. Notan. Collage of Agricultura, Food & Natural Resources, University of Missouri, Columbia, U.S.A.
[email protected] Ortega Corona. Instituto Nacional de
Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Ciudad Obregón, Sonora, México.
[email protected] Ortuño Sahagún. Departamento de Biología
Celular y Molecular, CUCBA. Zapopan, Jalisco. México. [email protected]
José Manuel Palma García. Universidad de Colima, México. [email protected]
Alfonso Peña Ramos. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP-Ags.), Aguas-calientes, México. [email protected]
Enrique Pimienta Barrios. Departamento de Producción Agrícola, CUCBA. Zapopan, Jalisco, México.
[email protected] Pratseval Algels. Centro de Tecnología de la Carne.
Monells (Girona) España. [email protected] Quesada Monge. Instituto Tecnológico de Costa
Rica. [email protected]é Luis Quintanar Stephano. Departamento de Fisiología
y Farmacología, Centro de Ciencias Básicas, Universidad Autónoma de Aguascalientes, México.
[email protected] Ramírez Juárez. Colegio de Posgraduados, Campus
Tecamachalco, Puebla, México. [email protected]
continúa en la tercera de forros

J. Antonio Rentería Flores. Centro Nacional de Investigación en Fisiología Animal. INIFAP. Ajuchitlan, Querétaro. México. [email protected]
Eduardo Ríos Jara. Departamento de Ecología, CUCBA. Zapopan, Jalisco, México. [email protected]
José de Jesús Sánchez González. Departamento de Producción Agrícola, CUCBA. Zapopan, Jalisco. México. [email protected]
Juan de Jesús Taylor Preciado. Departamento de Medicina Veterinaria, CUCBA. Zapopan, Jalisco. México. [email protected]
Joan Tibau. Centro de Control Porcino. Monells (Girona) España. [email protected]
Francisco José Trigo Tavera. División de Estudios de Posgrado e Investigación de la Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México. México. [email protected]
Enrique Trovo Diéguez. CIBNOR, Baja California, México. [email protected]
Raúl E. Vargas García. Departamento de Medicina Preventiva, Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de México. México. [email protected]
Mario Abel Vázquez García. Departamento de Producción Agrícola, CUCBA. Zapopan, Jalisco, México.
[email protected] Velásquez Martínez. Colegio de Posgraduados
de Chapingo, México. [email protected] Verdú Navarro. Departament de Biología Cel·lular,
de Fisiología i d’immunologia, Bellaterra, España. [email protected] Villalpando Ibarra. Organización Meteorológica
Mundial (OMM), Ginebra, Suiza. [email protected]
Comité Editorial (continuación)
DIRECTORIO DE LA UNIVERSIDAD DE GUADALAJARA
Carlos Jorge Briseño TorresRECTOR GENERAL
Gabriel Torres EspinozaVICERRECTOR EJECUTIVO
José Alfredo Peña RamosSECRETARIO GENERAL
Juan de Jesús Taylor PreciadoRECTOR DEL CENTRO UNIVERSITARIO DE CIENCIAS BIOLÓGICAS
Y AGROPECUARIAS
Enrique Pimienta BarriosSECRETARIO ACADÉMICO DEL CENTRO UNIVERSITARIO DE
CIENCIAS BIOLÓGICAS Y AGROPECUARIAS
Servando CarvajalCOORDINADOR DE INVESTIGACIÓN DEL CENTRO UNIVERSITARIO
DE CIENCIAS BIOLÓGICAS Y AGROPECUARIAS
scientia-CUCBA (ISSN: 1665-8493) es una publicación semestral (junio, diciembre) y está disponible mediante suscripción que puede hacerse a la Coordinación de Investigación del CUCBA, U. de G., km 15.5 Carretera Guadalajara a Nogales, Las Agujas, Zapopan, 45110, Jalisco, México. Tel.: (0133) 3777 1155; Fax.: (0133) 3777 1150. CERTIFICADO DE RESERVA DE DERECHOS AL USO EXCLUSIVO NÚMERO: 04-2003-101714124100-102
La opinión que se expresa en los artículos es responsabilidad de los autores. Se autoriza la reproducción total o parcial de los trabajos, siempre y cuando se cite la fuente y no sea con fi nes de lucro.
Auxiliar en la edición
MICHELLE ANN MERRITT WALTHER Coordinación General de Cooperación e Internacionalización. Unidad de Lenguas.Universidad de Guadalajara.
Diseño e impresión
TAGIT :: TECNOLOGÍA Y APLICACIONES GRÁFICASSaulo Cortés / José Manuel SánchezEnrique Díaz de León 514-2bGuadalajara 44170, Jalisco.TEL. (33) 3825•8528 FAX (33) 3825•[email protected] | tagit.idex.com.mx

scientia-CUCBA 9(2):81—93.2007
Efecto de la aplicación de agroquímicos y abonos orgánicos sobre la producción de frijol y la población microbiana del suelo
Manuel Morales Torres, Rogelio Lépiz Ildefonso, Diego R. González Eguiarte, Ricardo Nuño Romero, Luz Elena Claudio G., Patricia Zarazúa Villaseñor, Salvador Mena Munguía y Eduardo López AlcocerDepartamento de Producción Agrícola, Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de GuadalajaraKm 15.5 Carretera a Guadalajara-Nogales, Zapopan, C.P. 45101. Jalisco, México.Correo electrónico: [email protected]
Resumen: En Zapopan, Jalisco, en el ciclo agrícola de Primavera-Verano 2005, se realizó un experimento para estudiar la influencia de la aplicación de agroquímicos y abonos orgánicos al suelo, sobre el desarrollo y producción del frijol y sobre la población microbiana del suelo. El ensayo incluyó dos niveles de aplicación de insecticida, herbicida pre-emergente, herbicida pos-emergente, fertilizantes químicos, vermiabono, fermentado aeróbico, composta y micorrizas. En todos los casos, los niveles de aplicación fueron 0 (sin) y 1 (con); en total fueron 13 tratamientos, ubicados en un diseño experimental de bloques al azar con 4 repeticiones. Se registraron 6 variables de planta, 3 variables de suelo y 6 variables microbiológicas. Se realizaron análisis de varianza, prueba de medias y análisis de correlación. En las variables altura de planta, ramas en 10 plantas y rendimiento de grano, se detectaron diferencias significativas. En las variables de suelo, el efecto fue significativo para conductividad eléctrica, no así para materia orgánica, ni pH. En población microbiana del suelo, se encontraron diferencias en los dos muestreos de bacterias (floración y poscosecha) y en actinomicetes en muestreo de poscosecha; no se detectaron diferencias en hongos. Los resultados mostraron que la aplicación de insecticida al suelo, el uso de herbicidas y la aplicación de composta, afectaron positivamente a las variables altura de planta, ramas R9, y rendimiento de grano. La conductividad eléctrica se incrementó en el tratamiento con fertilización química. La población de bacterias resultó afectada por la aplicación de agroquímicos y mostró un claro descenso de siembra a cosecha, en tanto que los hongos no fueron afectados por los tratamientos. La población de actinomicetes fue significativamente diferente en la etapa de poscosecha y la mayor población se asoció con la aplicación de agroquímicos. Las variables altura de planta, biomasa R6, biomasa R8, ramas R9 y vainas R9, mostraron valores de correlación positivos con rendimiento de grano. Las variables de suelo materia orgánica y pH, mostraron valores de correlación positivos no significativos con rendimiento. Por su parte, la población bacteriana mostró correlación negativa con el rendimiento de grano y sus componentes.
Palabras clave: frijol, suelo, abonos orgánicos, agroquímicos, microorganismos del suelo, producción de grano.
Abstract: In Zapopan, Jalisco during the spring-summer agricultural cycle of 2005, an experiment was conducted in order to study the influence that the application of agrochemicals and organic manures to the soil has on the development and production of beans and on the microbic soil population. The test consisted in two levels of insecticide application, pre-emergent herbicide, post-emergent herbicide, chemical fertilizers, fermented aerobic casting, compost and mycorrhizae. In every instance the application levels were 0 (without) and 1 (with); there were 13 treatments in all, located in an experimental design of random blocks with 4 repetitions. Six plant variables were registered, as well as 3 soil and 3 microbiologic ones. An analysis of variance, a test of

82 Manuel Morales Torres et al.
scientia-CUCBA 9(2):81—93.2007
Introducción
EL CULTIVO DE FRIJOL
En México se siembran con frijol 2.25 mi-llones de hectáreas anuales, se producen 1.2 millones de toneladas y se importan 80 mil toneladas para cubrir las necesidades internas anuales de este grano básico. De la producción total, se destinan al consumo humano 1.12 millones de toneladas, dando como resultado un consumo anual de 11 ki-logramos por persona por año (CEA 2001; Lépiz et al. 2000).
Los cambios significativos ocurri-dos en la superficie cultivada de frijol en Jalisco, de 480,000 hectáreas en 1966 a 32,000 en 1999, reducción equivalente al 92%, han obedecido a factores tecnológi-cos y socioeconómicos. En maíz, cultivo con que se asociaba el frijol en un 91%, hoy día se utilizan altas densidades de población, herbicidas no selectivos para el frijol y se emplea maquinaria en las labo-res agrícolas, incluyendo la cosecha; estos componentes tecnológicos, han eliminado al frijol del sistema. Por otro lado, el siste-ma asociado maíz-frijol requiere de mayor número de jornales; la escasez de mano
means and a correlative analysis were performed. In the variables plant height, branches in 10 plants and grain yield, significant differences were found. In the soil variables, the effect for electric conductivity was significant, but not for organic matter nor for pH. In the microbic soil population differences were found in the two bacterial samples (blooming and post-crop) and in the post-crop Actomycetes sample; in fungi there were no differences. The results show that the application of insecticide to the soil, the use of herbicides and the application of compost positively affected the height variables of the plant, the branches R9 and the grain yield. Electric conductivity was increased in the treatment with chemical fertilization. The population of bacteria was affected by the application of agrochemicals and showed a marked decrease from sowing time to crop time, so much so that the fungi were not affected by the treatments. The population of Actinomycetes was significantly different in the post-crop stage, and a larger population was associated with the application of agrochemicals. The variables plant height, biomass R6, biomass R8, branches R9 and pods R9 show positive co-relational values with grain yield. The variables in organic soil matter and pH showed positive co-relational values of no significance and with yield. As for the bacterial population, it showed a negative correlation with the corn yield and its components.
Key words: bean, soil, organic fertilizers, agrochemicals, microorganisms of the soil, grain production.
de obra en el medio rural, se ha traducido igualmente en una reducción de la siembra de frijol en asociación con maíz (Lépiz et al. 2000).
Por lo que concierne a la agricultura en el municipio de Zapopan, de los años 60’s a la fecha, se ha adoptado el monocul-tivo de maíz con altas densidades de pobla-ción y aplicaciones irracionales de agroquí-micos, para mantener niveles satisfactorios de producción. Este cambio tecnológico ha afectado negativamente la fertilidad nativa del suelo, el contenido de materia orgáni-ca, las poblaciones de maleza y la actividad microbiana en los suelos, reduciendo de manera significativa la fertilidad de éstos.
VARIEDADES DE FRIJOL Y COMPONENTES DE RENDIMIENTO
Alemán et al. (1996) y Lépiz et al. (2007), coinciden en recomendar para la región Centro de Jalisco, las variedades Azufrado Tapatío, Alteño 2000, Bayo INIFAP y Flor de Mayo M-38. Los cultivares menciona-dos son de hábito de crecimiento indeter-minado postrado tipo III, propios para

Efecto de la aplicación de agroquímicos y abonos orgánicos sobre... 83
scientia-CUCBA 9(2):81—93.2007
siembras en unicultivo. Tiene un buen nivel de resistencia a las principales en-fermedades de la región como antracnosis [Colletotrichum lindemuthianum (Sacc. & Magn.) Br. & Cav.], roya [Uromyces appen-diculatus (Pers. & Unger.)], bacteriosis de halo (Pseudomonas syringae pv. phaseo-licola), bacteriosis común (Xanthomonas campestris pv. phaseoli), adaptación y un buen rendimiento, con producciones pro-medios de 2,000 kg/ha bajo condiciones de temporal y un buen manejo.
Los componentes del rendimiento, son características de la planta que se aso-cian positivamente con la producción de grano. En frijol se mencionan como com-ponentes a la biomasa, número de ramas, número de nudos y número de vainas por planta; también son componentes, granos por vaina y tamaño (peso) de grano (White e Izquierdo 1989 y Adams 1973).
ABONOS ORGÁNICOS
Los abonos orgánicos fueron la base de la fertilización durante muchos siglos, hasta la aparición de los fertilizantes químicos. Entre los desechos orgánicos más utiliza-dos, se mencionan al estiércol y el guano marino. En México, la aplicación de abo-nos orgánicos se remonta a la época de los aztecas y mayas, quienes utilizaban el pescado como fuente de fósforo y materia orgánica en las chinampas del Valle de México (Herrera et al. 2002).
La importancia que han merecido los abonos orgánicos, se debe entre otros motivos, al valor que tienen como mejora-dores de suelos, especialmente en aquellos con bajo contenido de materia orgánica, pobres en contenido de nutrientes y bajos en población microbiana. Esta situación es común en los suelos agrícolas sometidos a la aplicación continua de fertilizantes químicos, en suelos erosionados y com-pactados, con contenidos bajos de materia orgánica, como son los suelos del Valle de Zapopan. Los resultados obtenidos a lo largo del tiempo muestran que la aplica-ción prolongada de abonos orgánicos me-jora la estructura del suelo, incrementa la
cantidad y diversidad de microorganismos relacionados con la fertilidad, favorece la aireación, la infiltración y retención de la humedad, mejorando la fertilidad en gene-ral y favoreciendo directamente el desarro-llo y el rendimiento de los cultivos (Dick y McCoy 1993; Paino 1996).
De forma tradicional, durante años, los agricultores han reunido los desper-dicios orgánicos para transformarlos en abono para sus tierras, imitando y ace-lerando el proceso de fermentación que ocurre de manera natural en un suelo de un bosque; este proceso recibe el nombre de compostaje. El vermiabono, producto del incesante trabajo de ingestión y diges-tión de materiales orgánicos por parte de la lombriz de tierra, es otro abono orgánico de alto valor utilizado más recientemente; es soluble en agua, lo que permite preparar un abono liquido para poderlo mezclar en agua de riego (Reines 1998).
Por su parte, los fermentados aeró-bicos son abonos orgánicos líquidos con mucha energía equilibrada y en armonía mineral, preparados a base de estiércoles frescos disueltos en agua, que pueden ser enriquecidos con adiciones de leche, me-laza, cenizas, harinas de rocas minerales y sales minerales como sulfatos de magne-sio, zinc, cobre y otras. Sustituyen a los fer-tilizantes químicos industriales altamente solubles y fortalecen el equilibrio nutricio-nal a través de los ácidos orgánicos, las hormonas de crecimiento, antibióticos, vi-taminas, minerales, enzimas y coenzimas entre otros (Jairo 2005).
MICROORGANISMOS DEL SUELO
Kolmans (1995), estableció que los grupos de microorganismos presentes en el suelo, están integrados por bacterias, hongos y actinomicetes. Las bacterias comprenden el grupo más numeroso, variado y activo de microorganismos del suelo; son orga-nismos primitivos ubicados como proca-riotes unicelulares, carecen de membrana nuclear y de organelos, como mitocondrias y cloroplastos y se reproducen por bipar-tición, con rapidez asombrosa. Se encuen-

84 Manuel Morales Torres et al.
scientia-CUCBA 9(2):81—93.2007
tran ampliamente distribuidas en la natu-raleza, habitan en el agua, en el suelo, en la superficie o en el interior de las plantas y animales incluyendo al hombre. Pueden vivir en forma libre o en simbiosis con otros organismos, ya sea como patógenos, comensales o mutualistas (Madigan et al. 1999). Fisiológicamente este grupo pre-senta características muy variables; hay bacterias móviles e inmóviles, autótrofas fotosintéticas y quimiosintéticas y heteró-trofas (Atlas y Bartha 1997). Se desarrollan como parásitos y parcialmente en el suelo como saprófitas; la fertilidad del suelo está íntimamente relacionada con la cantidad de bacterias amonificantes y nitrifican-tes presentes, mismas que participan en la mineralización de la materia orgánica (Madigan et al. 1999).
Los hongos requieren de oxigeno para su metabolismo, por ello se encuen-tran y viven en suelos bien aireados, pre-sentan un sistema enzimático muy activo, característica que les permite degradar compuestos orgánicos muy resistentes como la lignina, principal fuente de hu-mus. Al carecer de clorofila, deben nutrirse de materia orgánica que degradan y mine-ralizan, poniéndola así a disposición de las plantas para su nutrición. Los hongos se ubican como heterótrofos eucariontes uni-celulares y pluricelulares con presencia de esporas y micelio; tienen hábitats muy di-versos; la mayoría son terrestres y habitan en el suelo, metabolizan materia orgánica muerta (saprófitos), desempeñando una actividad crucial en la mineralización del carbono y nitrógeno orgánicos.
Por su parte, los actinomicetes son un grupo intermedio entre las bacterias y los hongos; producen filamentos delga-dos, ramificados, que se desarrollan en un micelio, excepto el genero Actinomyces (Garassini 1962). Los actinomicetes parti-cipan en una gran cantidad de reacciones bioquímicas, por lo que degradan muchos compuestos orgánicos, segregan antibióti-cos que benefician la presencia de hongos y la formación de humus. Se desarrollan de manera más lenta que la mayoría de hon-
gos y bacterias, característica que indica su incapacidad como competidores efectivos; llegan a predominar cuando los nutrientes comienzan a ser limitantes y la presión de los competidores más efectivos disminuye (Sykes y Skinne, 1999).
Por otra parte, se conoce con el nom-bre de micorriza a la asociación mutualista entre las raíces de la mayoría de las plantas y diversos hongos del suelo. Se trata de una simbiosis presente en casi todas las plan-tas; el hongo coloniza la raíz de la planta proporcionándole nutrientes y agua, por medio de la extensa red de hifas, mientras que la planta suministra al hongo sustra-tos energéticos y carbohidratos (Madigan et al. 1999). El efecto más importante que producen las micorrizas en las plantas es un incremento en la absorción de nutrien-tes del suelo, que se traduce en un mayor crecimiento y desarrollo de las mismas. La expansión del micelio externo por el suelo rizosférico es la causa principal de este efecto, permitiendo la captación de los nu-trientes más allá de la zona de agotamiento que se crea alrededor de las raíces, por la propia absorción de las plantas (Brougher y Malajczuk 1990).
FERTILIZANTES QUÍMICOS
En cuanto a los nutrimentos proporcio-nados como fertilizantes, los resultados de numerosas investigaciones y la vasta experiencia de los productores agrícolas, señalan que los diferentes cultivos respon-den positivamente a la aplicación de fertili-zantes químicos a base de N, P y K.
Un suministro adecuado de nitróge-no, elemento que participa en la compo-sición de las más importantes sustancias orgánicas, produce un rápido y vigoroso crecimiento de la planta, color verde inten-so en las hojas, aumento del contenido de proteínas y un aumento en la producción de hojas, frutos y semillas (Domínguez 1993). Desde el punto de vista de la ferti-lidad del suelo, las formas iónicas NH4
+, NO2
- y NO3- son las de mayor importancia:
el oxido nitroso y el oxido nítrico también son importantes, porque representan for-

Efecto de la aplicación de agroquímicos y abonos orgánicos sobre... 85
scientia-CUCBA 9(2):81—93.2007
mas de nitrógeno que se pierden a través de la desnitrificación (Chandra y Bollen 1980).
Por lo que concierne al fósforo, en la mayoría de las plantas se encuentra en me-nores cantidades en relación a nitrógeno y potasio. Las plantas absorben gran parte del fósforo en forma del ión primario or-tofosfato H2PO4
- y en segundo término del HPO4
2-. Las cantidades relativas de absor-ción de los dos iones, están afectadas por el potencial de Hidrógeno (pH) del medio que rodea a las raíces. Los valores bajos del pH incrementan la absorción del ión H2PO4
-, mientras que los valores más altos del pH incrementan la absorción de la forma HPO4
2- (Chai y Caldwell 1979). Un sumi-nistro adecuado de fósforo en las primeras etapas de la vida de la planta, es importan-te para evitar el retraso del crecimiento de las partes reproductivas; activa la madurez de las plantas, se asocia con una solidez de los tallos y aumenta la resistencia a las en-fermedades (Bixby et al. 1974).
De acuerdo con Alemán et al. (1996) y Lépiz et al. (2007), el frijol de temporal en la región centro de Jalisco muestra respuesta positiva a las aplicaciones de nitrógeno y fósforo en cantidades que va-rían de 40 a 50 kg por hectárea de ambos elementos. Para sitios de baja precipitación y suelos delgados, recomiendan el trata-miento 30-30-0, en tanto que para sitios de buena precipitación y suelos profundos de mediana a baja fertilidad, se debe apli-car el tratamiento 50-50-0 de N y P2O5, respectivamente. Todo el fertilizante debe aplicarse en la siembra y cubrirse con una capa de suelo, antes de la siembra.
INSECTICIDAS
El empleo de insecticidas en la agricultura moderna, es una práctica cotidiana y en la mayoría de los casos necesaria. En la tota-lidad de las hortalizas y de algunas legu-minosas como el frijol, si no se controlan las plagas insectiles en el cultivo, los daños pueden acabar con la cosecha. Por la razón anterior, los manuales de producción de cultivos incluyen el capítulo de plagas y su
control. En este sentido, Alemán et al. (op. cit.) y Lépiz et al. (op. cit.), dieron a cono-cer las principales plagas del frijol y de su control, en la región centro de Jalisco.
HERBICIDAS
Debido a la buena disponibilidad de hume-dad y suelos profundos en la región cen-tro de Jalisco, las malezas herbáceas son abundantes y competitivas con el frijol. Para evitar daños al cultivo, deben remi-tirse por lo menos durante los primeros 40 días después de la siembra. En siembras mayores de una hectárea, se hace impres-cindible el uso de herbicidas. En pre-emer-gencia, pueden utilizarse las mezclas a base de Lazo + Afalón o Prowl + Bladex, a razón de 1.5 + 0.750 litros por hectárea. De post-emergencia, se recomienda la mezcla Flex + Fusilade 0.5 litros de cada uno por hectárea (Lépiz et al., 2007).
SISTEMA DE PRODUCCIÓN EN LA REGIÓN DE ZAPOPAN
La agricultura en la región Centro de Jalisco a partir de los años 60, adoptó el monocultivo de maíz, sistema que depende de la utilización de altas densidades de po-blación, de la aplicación de dosis irraciona-les de fertilizantes, uso intensivo de herbi-cidas e insecticidas, para mantener niveles satisfactorios de producción de grano. Este cambio tecnológico afectó de manera ne-gativa la fertilidad nativa del suelo, el con-tenido de materia orgánica, las poblaciones de maleza y la actividad microbiana en los suelos, reduciendo de manera significativa la fertilidad de los suelos.
Si se considera la problemática se-ñalada y sobre todo la relacionada con la degradación de los suelos en la región de Zapopan, se planteó la realización de la presente investigación, con los objetivos específicos de conocer la influencia de los agroquímicos y abonos orgánicos aplicados al suelo, sobre el desarrollo y producción de frijol y sus efectos sobre la población microbiana del mismo.

86 Manuel Morales Torres et al.
scientia-CUCBA 9(2):81—93.2007
Materiales y métodos
El ensayo se estableció en el Municipio de Zapopan, Jalisco, durante el ciclo Primavera-Verano 2005. Se incluyeron 13 tratamientos en un diseño de bloques al azar con cuatro repeticiones, para estu-diar el efecto de la aplicación al suelo de insecticidas, herbicidas pre-emergentes, herbicidas post-emergentes, fertilizantes químicos, vermiabono, fermentado aeróbi-co, composta y micorrizas. En todos los ca-sos, los niveles de aplicación fueron 0 (sin) y 1(con). La unidad experimental fue de cinco surcos de 5 metros a 70 cm de sepa-ración, tomando como parcela útil los tres surcos centrales. Para la siembra se abrió una raya profunda en el lomo del surco, se aplicaron los insumos correspondientes a cada parcela, se cubrieron con una capa delgada de suelo y se sembró. Se utilizó la variedad de frijol Azufrado Tapatío, depo-sitando 100 semillas por metro lineal de surco.
Se evaluaron seis variables de planta, tres de suelo y seis microbiológicas. Las variables de suelo y de microorganismos se cuantificaron en las repeticiones 1 y 3. Para la estimación de la población microbiana (E.P.M.) utilizando el método de vaciado en placa con inoculación por superficie, se partió de 10 g de suelo y se realizaron nueve diluciones, las cuales se sembraron en los medios de cultivo agar nutritivo para bacterias, PDA para hongos y Czapec Dox para actinomicetes; las cajas de Petri se incubaron a 37 grados centígrados du-rante 24 horas, 72 horas y 6 días respecti-vamente; luego se realizó la cuantificación de colonias (UFC), considerando colonias individuales a todas aquellas que distaron de las colonias próximas, al menos un diá-metro de la colonia más pequeña; el valor obtenido se multiplicó por el factor de dilu-ción y se dividió entre la cantidad de suelo usado. Para expresar la cantidad en menor número de dígitos, se utilizaron exponen-tes. Para bacterias se utilizó el E.P.M. x g suelo x 10-16; para hongos y actinomicetes E.P.M. x g suelo 10-12. El método de la di-
lución en placa con inoculación por super-ficie, sólo determina células microbianas heterótrofas aeróbicos viables.
Resultados y discusión
ANÁLISIS DE VARIANZA
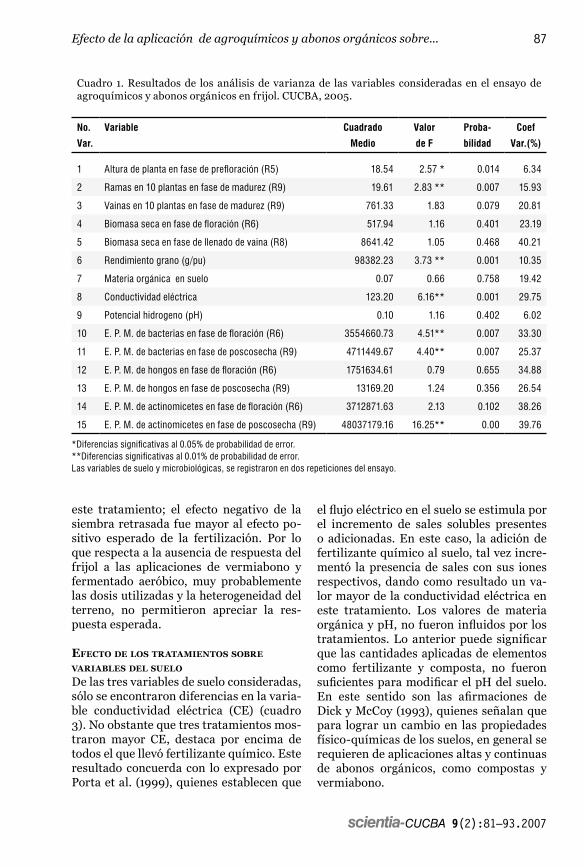
Los resultados de los análisis de varianza practicados en cada una de las 15 varia-bles, se muestran en el cuadro 1. En las va-riables de planta se detectaron diferencias en altura, número de ramas y rendimiento de grano. En las variables de suelo, se en-contraron diferencias significativas, solo para conductividad eléctrica (CE). En la estimación de la población microbiana (EPM), el análisis detectó diferencias alta-mente significativas en bacterias en los dos muestreos y en actinomicetes, en el segun-do muestreo; en hongos no hubo efecto de los tratamientos.
EFECTO DE LOS TRATAMIENTOS SOBRE LAS VARIABLES DE PLANTA
Las variables ramas en planta y rendi-miento de grano, mostraron una respuesta similar y positiva a las aplicaciones de in-secticida al suelo, herbicida pre-emergente y composta como factor común (cuadro 2). Los resultados obtenidos en las variables mencionadas, mostraron el comporta-miento esperado; es decir, la aplicación de insecticida al suelo, el uso de herbicidas y la aplicación de composta, se expresan en un mayor número de ramas y mayor rendimiento de grano. Concuerda con lo expresado por Dick y McCoy (1993) y Paino (1996) entre otros, quienes señalan que la aplicación de compostas maduras mejora la fertilidad del suelo. Las aplicaciones de insecticida y herbicidas, contribuyeron a un mejor control de plagas y maleza, lo que dio como resultado un mejor desarrollo del frijol y mayor rendimiento (Alemán et al. op. cit.; y Lépiz et al. op. cit.).
La ausencia de respuesta del frijol a la fertilización química, contraria a lo se-ñalado en la literatura (Alemán et al. op. cit.; y Lépiz et al. op. cit.), se explica por la siembra tardía (resiembra) del frijol en
Efecto de la aplicación de agroquímicos y abonos orgánicos sobre... 87
scientia-CUCBA 9(2):81—93.2007
Cuadro 1. Resultados de los análisis de varianza de las variables consideradas en el ensayo de agroquímicos y abonos orgánicos en frijol. CUCBA, 2005.
No.
Var.
Variable Cuadrado
Medio
Valor
de F
Proba-
bilidad
Coef
Var.(%)
1 Altura de planta en fase de prefloración (R5) 18.54 2.57 * 0.014 6.34
2 Ramas en 10 plantas en fase de madurez (R9) 19.61 2.83 ** 0.007 15.93
3 Vainas en 10 plantas en fase de madurez (R9) 761.33 1.83 0.079 20.81
4 Biomasa seca en fase de floración (R6) 517.94 1.16 0.401 23.19
5 Biomasa seca en fase de llenado de vaina (R8) 8641.42 1.05 0.468 40.21
6 Rendimiento grano (g/pu) 98382.23 3.73 ** 0.001 10.35
7 Materia orgánica en suelo 0.07 0.66 0.758 19.42
8 Conductividad eléctrica 123.20 6.16** 0.001 29.75
9 Potencial hidrogeno (pH) 0.10 1.16 0.402 6.02
10 E. P. M. de bacterias en fase de floración (R6) 3554660.73 4.51** 0.007 33.30
11 E. P. M. de bacterias en fase de poscosecha (R9) 4711449.67 4.40** 0.007 25.37
12 E. P. M. de hongos en fase de floración (R6) 1751634.61 0.79 0.655 34.88
13 E. P. M. de hongos en fase de poscosecha (R9) 13169.20 1.24 0.356 26.54
14 E. P. M. de actinomicetes en fase de floración (R6) 3712871.63 2.13 0.102 38.26
15 E. P. M. de actinomicetes en fase de poscosecha (R9) 48037179.16 16.25** 0.00 39.76
*Diferencias significativas al 0.05% de probabilidad de error.**Diferencias significativas al 0.01% de probabilidad de error.Las variables de suelo y microbiológicas, se registraron en dos repeticiones del ensayo.
este tratamiento; el efecto negativo de la siembra retrasada fue mayor al efecto po-sitivo esperado de la fertilización. Por lo que respecta a la ausencia de respuesta del frijol a las aplicaciones de vermiabono y fermentado aeróbico, muy probablemente las dosis utilizadas y la heterogeneidad del terreno, no permitieron apreciar la res-puesta esperada.
EFECTO DE LOS TRATAMIENTOS SOBRE VARIABLES DEL SUELO
De las tres variables de suelo consideradas, sólo se encontraron diferencias en la varia-ble conductividad eléctrica (CE) (cuadro 3). No obstante que tres tratamientos mos-traron mayor CE, destaca por encima de todos el que llevó fertilizante químico. Este resultado concuerda con lo expresado por Porta et al. (1999), quienes establecen que
el flujo eléctrico en el suelo se estimula por el incremento de sales solubles presentes o adicionadas. En este caso, la adición de fertilizante químico al suelo, tal vez incre-mentó la presencia de sales con sus iones respectivos, dando como resultado un va-lor mayor de la conductividad eléctrica en este tratamiento. Los valores de materia orgánica y pH, no fueron influidos por los tratamientos. Lo anterior puede significar que las cantidades aplicadas de elementos como fertilizante y composta, no fueron suficientes para modificar el pH del suelo. En este sentido son las afirmaciones de Dick y McCoy (1993), quienes señalan que para lograr un cambio en las propiedades físico-químicas de los suelos, en general se requieren de aplicaciones altas y continuas de abonos orgánicos, como compostas y vermiabono.

88 Manuel Morales Torres et al.
scientia-CUCBA 9(2):81—93.2007
EFECTO DE LOS TRATAMIENTOS SOBRE LAS VARIABLES MICROBIOLÓGICAS
Según se muestra en el cuadro 1, se detec-taron diferencias altamente significativas (p < 0.05) en la estimación de la pobla-ción microbiana en bacterias en los dos muestreos (floración y poscosecha) y en el segundo muestreo (poscosecha) de actino-micetes.
En bacterias se formaron seis grupos de tratamientos estadísticamente iguales para el primer muestreo y cinco para el segundo. En ambos casos se aprecia una clara tendencia de menor número de bacte-rias en los tratamientos con agroquímicos. Este resultado sugiere que la aplicación de
insecticida, herbicidas y fertilizante quí-mico, pudo afectar de manera negativa a la población bacteriana del suelo (cuadro 4). Sobre este particular Madigan et al. (1998), sostienen que algunos factores am-bientales como los fertilizantes químicos y pesticidas, afectan el desarrollo micro-biano, pudiendo incluso causar su muerte. Además, la aplicación de vermiabono, fer-mentado aeróbico y micorrizas, muestran una asociación positiva con el número de bacterias.
Por otra parte, en la figura 1 se ob-serva un comportamiento descendente del número promedio de bacterias en el ensayo, de 17,000 a 4,000 E.P.M/g suelo
Cuadro 2. Resultados obtenidos en las variables registradas en la planta de frijol en el ensayo de agroquímicos y abonos orgánicos en frijol. CUCBA, 2005.
No. Trat.
TratamientosIS-HA-HD-FQ-VA-
FA-CP-MR1
AlturaPlanta (cm)
Ramas 10 Plantas
Vainas 10
Plantas
Biomasa R6 g/10 plantas
Biomasa R8 g/10 plantas
Rendimientode Grano
(g/pu)
5 0-1-0-0-0-0-1-0 41.15 17.75 112.3 77.50 276.50 1779
3 0-0-1-0-0-0-1-0 41.05 16.25 97.50 97.50 223.50 1762
11 0-0-0-0-0-0-0-0 42.75 16.75 103.8 100.00 240.50 1759
2 1-0-0-0-0-0-1-0 44.25 17.25 108.3 77.50 196.50 1657
6 1-1-0-0-0-0-1-0 44.50 20.75 116.8 102.50 255.50 1620
4 1-0-1-0-0-0-1-0 41.05 16.75 99.50 87.50 204.50 1606
7 0-1-1-0-0-0-1-0 41.60 15.75 105.0 107.50 233.00 1605
10 0-0-0-0-1-0-0-0 42.25 12.25 68.25 95.00 172.00 1557
1 0-0-0-0-0-0-1-0 44.60 16.50 105.3 100.00 370.50 1541
8 1-1-1-0-0-0-1-0 43.40 20.00 103.5 115.00 292.00 1434
13 0-0-0-0-0-0-0-1 44.20 14.75 89.25 87.50 212.00 1418
12 0-0-0-0-0-1-0-0 43.30 14.50 84.25 85.00 135.00 1366
9 0-0-0-1-0-0-0-0 36.75 15.75 80.50 52.50 125.00 1284
Promedio 42.37 16.54 98.00 91.15 225.88 1568
C. V. (%) 6.34 15.93 20.81 23.19 48.30 10.35
F 0.05 2.57 * 2.83 ** 1.83 1.16 1.05 3.73 **
DMS 0.05 3.85 3.78 29.24 49.59 202.05 232
1IS, insecticida al suelo; HA, herbicida pre-emergente; HD, herbicida post-emergente; FQ, fertilizante químico; VA, vermiabono; FA, fermentado aerobio; CP, composta; MR, micorriza.*Diferencias significativas al 0.05% de probabilidad de error.**Diferencias significativas al 0.01% de probabilidad de error.

Efecto de la aplicación de agroquímicos y abonos orgánicos sobre... 89
scientia-CUCBA 9(2):81—93.2007
Cuadro 3. Resultados obtenidos en las variables de suelo, en el ensayo de agroquímicos y abonos orgánicos en frijol. CUCBA, 2005.
No. Trat.
TratamientosIS-HA-HD-FQ-VA-FA-CP-MR*
MateriaOrgánica
(%)
ConductividadEléctrica
(mmhos/cm2)
Potencial Hidrogeno(pH)
5 0-1-0-0-0-0-1-0 1.950 15.00 4.755
2 1-0-0-0-0-0-1-0 1.910 17.00 4.945
4 1-0-1-0-0-0-1-0 1.850 15.50 5.195
13 0-0-0-0-0-0-0-1 1.815 12.00 5.380
7 0-1-1-0-0-0-1-0 1.765 11.00 5.295
10 0-0-0-0-1-0-0-0 1.720 6.00 5.065
8 1-1-1-0-0-0-1-0 1.675 14.50 4.715
6 1-1-0-0-0-0-1-0 1.585 13.50 5.405
12 0-0-0-0-0-1-0-0 1.585 15.00 5.080
11 0-0-0-0-0-0-0-0 1.585 11.00 5.210
1 0-0-0-0-0-0-1-0 1.490 12.50 5.285
3 0-0-1-0-0-0-1-0 1.480 13.00 5.110
9 0-0-0-1-0-0-0-0 1.315 39.50 4.790
Promedio 1.67 15.01 5.09
C. V. (%) 19.42 29.75 6.02
F 0.05 0.66 6.16** 0.00
DMS 0.05 0.58 24.16 0.72
1IS, insecticida al suelo; HA, herbicida pre-emergente; HD, herbicida post-emergente; FQ, fertilizante químico; VA, vermiabono; FA, fermentado aerobio; CP, composta; MR, micorriza.**Diferencias significativas al 0.01% de probabilidad de error.
��������������
�����������������������
������
������
������
������
������
�����
�����
�����
�����
�������������������������������������������
����������������������������
Figura 1. Comportamiento promedio de los microorganis-mos en todos los tratamientos y muestreos. CUCBA, 2005.

90 Manuel Morales Torres et al.
scientia-CUCBA 9(2):81—93.2007
x 10-16, desde el momento de presiem-bra (29/07/05), a la época de poscosecha (13/12/05). El descenso observado podría atribuirse a la combinación de menor humedad disponible en el suelo y a la pre-sencia de temperaturas menores, hacia el final del ciclo y estación de crecimiento (Madigan et al. 1998).
En relación a la variable población de actinomicetes, en los dos muestreos el tratamiento 9 que llevó fertilizante quími-co, se ubicó como primero en las tablas de comparación de promedios (cuadro 4). Los tratamientos 4, 8 y 2, del grupo de mayor población de actinomicetes del muestreo de poscosecha, llevaron insecticida y dos de ellos herbicida. Este resultado permite
Cuadro 4. Resultados obtenidos en las variables de la población microbiana estimada por gramo de suelo, en el ensayo de agroquímicos y abonos orgánicos en frijol. CUCBA, 2005.
No. Trat.
Tratamientos IS-HA-HD-FQ-VA-
FA-CP-MR1
Bacterias R6
(x 10-16)
Bacterias R9
(x 10-16)
Hongos R6 (x 10-12)
Hongos R9 (x 10-12)
Actinomi-cetos R6 (x 10-12)
Actinomi-cetos R9 (x 10-12)
12 0-0-0-0-0-0-0-1 19100 6050 4850 464 4675 62
11 0-0-0-0-0-0-0-0 15940 3283 4400 514 4013 6135
10 0-0-0-0-1-0-0-0 12960 7425 3925 310 4513 370
7 0-1-1-0-0-0-1-0 11800 3535 3600 386 3000 120
1 0-0-0-0-0-0-1-0 10650 3250 4775 319 5350 60
13 0-0-0-0-0-1-0-0 9730 5500 5150 535 4550 6433
4 1-0-1-0-0-0-1-0 7905 4293 5250 283 2250 112920
3 0-0-1-0-0-0-1-0 7713 4293 2400 394 2250 172
9 0-0-0-1-0-0-0-0 7238 4770 5550 332 5500 11910
8 1-1-1-0-0-0-1-0 5435 2020 2750 385 2525 10250
6 1-1-0-0-0-0-1-0 4640 2525 4475 362 1500 542
2 1-0-0-0-0-0-1-0 4363 2778 3975 307 2768 8808
5 0-1-0-0-0-0-1-0 3833 3283 4425 448 2000 75
Promedio 9330 4077 4271 388 3453 4325
C. V. (%) 33.30 25.37 34.88 26.54 38.26 39.76
F 0.05 4.51** 4.40** 0.79 1.24 2.13 16.25**
DMS 0.05 6770 2254 3246 224 2878 3746
1IS, insecticida al suelo; HA, herbicida pre-emergente; HD, herbicida post-emergente; FQ, fertilizante químico; VA, vermiabono; FA, fermentado aerobio; CP, composta; MR, micorriza.**Diferencias significativas al 0.01% de probabilidad de error.
inferir, que la aplicación de agroquímicos al suelo favoreció de alguna manera la po-blación de actinomicetes, resultado inver-so al comportamiento general observado en bacterias. Sobre este tema, la literatura señala que la población de actinomicetes se incrementa al reducirse la competencia por bacterias y hongos, en especial cuando las condiciones ambientales son adversas (Madigan et al. 1998; Sykes y Skinne 1999). En consecuencia, puede afirmarse que el incremento observado en la población de actinomicetes en los tratamientos que llevaron agroquímicos, fue por la menor población y competencia de bacterias y no tanto por el efecto benéfico de los agroquí-micos.

Efecto de la aplicación de agroquímicos y abonos orgánicos sobre... 91
scientia-CUCBA 9(2):81—93.2007
CORRELACIONES
El cuadro 5 muestra el resultado de la correlación múltiple ente las 15 variables consideradas en el trabajo. En total hubo 18 correlaciones con valores significativos. Con rendimiento de grano, la variable de planta agronómicamente más importante, mostraron valores de correlación positivos altura, biomasa R6, biomasa R8, ramas R9 y vainas R9, dando valores significativos las correlaciones de biomasa R8 y vainas R9. Este resultado está de acuerdo con lo esperado y con lo reportado en la literatura sobre componentes de rendimiento, pues en todos los casos, una mayor producción de biomasa, ramas y vainas, se asocia con un mayor rendimiento (Adams 1993; White e Izquierdo 1991).
Las variables de suelo, materia orgá-nica (MO) y pH, mostraron valores positi-vos no significativos con rendimiento. Es de esperar que la adición de MO muestre una tendencia a asociarse con rendimiento y que al subir los niveles de pH en los sue-los ácidos del sitio del ensayo (promedios de 5.0), también el frijol tienda a subir los rendimientos, según lo refieren Madigan et al. (1998)
En relación a las correlaciones de los microorganismos en el suelo con las varia-bles de planta, en todos los casos fueron negativas. Por ejemplo, en bacterias R9, se observó una correlación negativa y signifi-cativa con biomasa R9, ramas R9 y vainas R9, debido al descenso pronunciado de las bacterias al final del ciclo del cultivo (figura 1). Hubo ausencia de correlación de los mi-croorganismos con las variables del suelo MO, pH CE; este resultado se explica, posi-blemente, por los bajos niveles aplicados de los abonos orgánicos, como vermiabono, composta y fermentado aerobio, niveles que no afectaron las propiedades físicas y químicas del suelo.
Conclusiones
De acuerdo con los resultados obtenidos en el ensayo, con los factores y niveles es-tudiados, los materiales empleados y bajo las condiciones en que la investigación se realizó, se concluye lo siguiente: a) la aplicación de insecticida al suelo,
el uso de herbicidas y la aplicación de composta, afectaron positivamente a las
Cuadro 5. Resultados de la correlación múltiple entre 15 variables en el ensayo de agroquímicos y abonos orgánicos en frijol. CUCBA. 2005.
V1 V2 V3 V4 V5 V6 V7 V8 V9 V10 V11 V12 V13 V14 V15V1 1.000V2 0.604 1.000V3 0.490 0.592 1.000V4 0.225 0.288 0.517 1.000V5 0.371 0.341 0.664 0.828 1.000V6 0.348 0.329 0.546 0.155 0.553 1.000V7 0.338 0.042 0.086 0.041 0.253 0.260 1.000V8 -0.711 -0.768 -0.410 0.101 -0.164 -0.588 -0.377 1.000V9 0.475 0.315 0.110 -0.140 0.088 0.235 -0.008 -0.411 1.000V10 0.087 0.112 -0.307 -0.655 -0.496 -0.158 -0.199 -0.272 0.326 1.000V11 -0.228 -0.310 -0.639 -0.901 -0.907 -0.308 -0.087 -0.066 0.164 0.581 1.000V12 -0.166 -0.611 -0.271 -0.218 -0.200 -0.281 0.142 0.418 0.251 0.209 0.281 1.000V13 0.193 0.106 -0.011 -0.064 0.039 0.180 0.047 -0.212 0.192 0.371 0.068 0.033 1.000V14 -0.146 -0.344 -0.237 -0.658 -0.668 -0.445 -0.343 0.279 0.065 0.591 0.544 0.478 0.104 1.000V15 0.212 -0.024 0.436 0.308 0.512 0.534 0.014 -0.074 0.087 -0.453 -0.393 -0.115 -0.493 -0.367 1.000
1, Altura de Planta en Floración (R6); 2, Biomasa Seca en Floración (R6); 3, Biomasa Seca en Llenado de Vaina (R8); 4, Ramas de 10 Plantas en Madurez (R9); 5, Vainas de 10 plantas en Madurez (R9); 6, Rendimiento de Grano (g/pu); 7, Materia Orgánica en el suelo (%); 8, Conductividad Eléctrica en el suelo (mmhos/cm2); 9, Potencial Hidrogeno en el suelo (pH); 10, Bacterias M1 (E.P.M./g suelo x10-16); 11, Bacterias M 2 (E.P.M./g suelo x 10-16); 12, Hongos M1 (E.P.M./g suelo x 10-12); 13, Hongos M2 (E.P.M./g suelo x 10-12); 14, Actinomicetes M1 (E.P.M./g suelo x 10-12); 15, Actinomicetes M2 (E.P.M./g suelo x 10-12).

92 Manuel Morales Torres et al.
scientia-CUCBA 9(2):81—93.2007
variables ramas en madurez (R9) y ren-dimiento de grano del frijol;
b) los tratamientos estudiados, no influye-ron de manera significativa el contenido de materia orgánica, ni los valores de pH en el suelo;
c) la población de bacterias resultó afecta-da por la aplicación de agroquímicos y mostró un claro descenso de siembra a poscosecha;
d) los hongos no mostraron efecto de los tratamientos y la población de actino-micetes mostró asociación positiva con la aplicación de agroquímicos al suelo; d) el rendimiento de grano mostró va-lores de correlación positivos con las variables altura de planta, biomasa R6, biomasa R8, ramas R9 y vainas R9;
e) la población bacteriana mostró en gene-ral, una tendencia de correlación nega-tiva con el rendimiento de grano y sus componentes.
Literatura citada
Adams, W. 1973. Plant architecture and phy-siological efficiency in the field bean. In: CIAT (Ed.). Potentials of field beans and other food legumes in Latin America. CIAT, Cali, Colombia. Pp. 37–67.
Alemán, V., S. Núñez, H. Flores, P. Alemán y J. Aceves. 1996. Guía para producir fri-jol en los Altos y Centro de Jalisco. Agenda Técnica No. 2. Campo Experimental de Los Altos de Jalisco. CIRPAC, INIFAP. 36 pp.
Atlas R., M. and R. Bartha. 1997. Microbial Ecology. Fundamentals and applications. Benjamin/Cummings Science Publishing. U. S. A. 567 pp.
Bixby, D., D. Rucker and S. Tisdale. 1974. Phosphoric fertilizers, properties and pro-cesses. Tech. Bull. No. 8 Washington, D.C. 151 p.
Brougher, N. and N. Malajczuk. 1990. Effect of high soil moisture on formation of Ectomycorrhizas and gtrowth of karri (Eucalyptus diversicolor). Seedlings ino-culated with Descolea maculata, Pisolithus
tinctorius and Laccaria laccata. Canadian Journal of Soil Science 74: 367–386.
CEA. 2001. Situación actual y perspectivas de la producción de frijol en México. Centro de Estadística Agropecuaria y Pesquera. SAGARPA. México, D. F. 61 pp.
Chai, M. and A. Caldwel. 1979. Forms of Phosphorus and Fixation in Soils. Soil Science Society of America (SSSA). Washington, USA. 23-75 pp.
Chandra, P. and W. Bollen. 1980. Effects of gribel on nitrification and sulfur oxida-tion in different Oregon soils apply. Journal Microbiol. Vol. :31–40.
Dick, W. and E. McCoy. 1993. Enhancing soil fertility by addition of compost. In: Science and Engineering of Composting: Design, environmental, microbiological and utili-zation aspects. The Ohio State University. 327 pp.
Domínguez, A. 1993. Abonos Orgánicos. Ministerio de Agricultura. Madrid, España. 105 pp.
Garassini, L. 1962. El suelo y su microflora. Universidad Central de Venezuela. Facultad de Agronomía. Maracay, Venezuela. 225 p.
Herrera, L., I. Fernández y N. González. 2002. Fertilización a partir de materiales orgánicos en una rotación de cultivos. In: III Encuentro Nacional de Agricultura Orgánica. Programas y Resúmenes. Universidad Central de las Villas. Cuba. pp. 4–12.
Jairo, R. 2005. Agricultura orgánica, bio-fertilizantes preparados y fermentados a base de mierda de vaca. Fundación Juquira Candiru, Río de Janeiro, Brasil. 96 pp.
Kolmans, E. 1995. La Agricultura como base para un desarrollo rural sustenta-ble. Centro Americano sobre Agricultura Orgánica. Acuerdo Bilateral de Desarrollo Sustentable Costa Rica-Holanda. 215 pp.
Lépiz I., R., E. López, S. Núñez, I. J. González, A. Ledesma y S. Herrera. 2000. Perspectivas del frijol en el Occidente de México. U. de G., INIFAP. Documento de circulación interna. 40 pp.
Lépiz I., R., S. Sánchez, E. López, A. González y S. Núñez. 2007. El cultivo de frijol en la regiones Centro y Sur de Jalisco. Tecnología para altos rendimientos. U. de G., INIFAP. En prensa. 42 pp.
Madigan, T., J. Martinko y J. Parker. 1999. Brock, biología de los microorganis-mos. Prentice Hall Iberia. Madrid, España. 1664 pp.

Efecto de la aplicación de agroquímicos y abonos orgánicos sobre... 93
scientia-CUCBA 9(2):81—93.2007
Paino, V. 1996. Municipal tropical compost: effects on crops and soil properties. In: Compost Science & Utilization. Springfield. 4(2): 341 pp.
Reines, M. 1998. Lombricultura, alternativa del desarrollo sustentable. Universidad de Guadalajara, CUCBA. México. 67 pp.
Sykes, G. and F. Skinne. 1999. Magazine Actinomycetales. Academic Press, Univer-sity of New York. USA. 123: 45–62.
White, J. y J. Izquierdo. 1989. FRIJOL: Fisiología del potencial de rendimiento y la tolerancia al estrés. CIAT-FAO. Santiago, Chile. 91 pp.
FECHA EFECTIVA DE PUBLICACIÓN 29 DE DICIEMBRE DE 2007

94
scientia-CUCBA 9(2):94.2007

scientia-CUCBA 9(2):95—103.2007
Sistemas silvopastoriles para una reconversión ambiental de la ganadería en la cuenca del río San Pedro en Camagüey, Cuba
Zoe G. Acosta GutiérrezCentro de Investigaciones de Medio Ambiente de Camagüey. Cisneros No. 105 (altos) e/ Ángel y Pobre. C.P. 70 100 Camagüey, Cuba. Telefax: (53) (32) 296349.Correo electrónico: [email protected]
Resumen: Se determinaron las posibles especies arbóreas a establecer en sistemas silvopastoriles como vía de reforestación de las diferentes entidades que desarrollan la ganadería en la cuenca hidrográfica del río San Pedro, Camagüey, Cuba. Para ello se utilizó un sistema de información geográfica (SIG) el cual asistió al cálculo de las áreas, a la determinación y ubicación de los diferentes tipos de suelo así como a la precisión del comportamiento de las principales variables climáticas. En las áreas ganaderas a reforestar predominan los suelos que clasifican como Inceptisoles los cuales ocupan un 59% del área total, seguidos por los Mollisoles con 143 km2 (22%), los Entisoles con 95 km2 (14%) y los Vertisoles con sólo 31 km2 (5%). El índice climático conocido como razón de evapotranspiración (E/E0), se comporta en un rango adecuado para el establecimiento de sistemas con árboles y pastos en las áreas ganaderas de la cuenca. Se presenta un listado de especies afines a los suelos presentes, las cuales se deberían considerar al momento de planificar la reforestación de las áreas bajo estudio, Se evalúa con vistas a un mejoramiento de la gestión ambiental y productiva.
Palabras claves: sistemas silvopastoriles, ganadería sostenible.
Abstract: The tree species able to be used in the silvopastoral systems in the reforestation of cattle areas of San Pedro watershed in Camagüey, Cuba were determined. A geographic information system (GIS) was used to calculate the areas of different zones, to determine and locate different soils, as well as to identify the behavior of the main climatic variables. In cattle areas selected to reforest, the soils classified as Incepetisols prevail and cover the 59% of the total area, followed by Mollisols with 142 km2 (22%), Entisols with 95 km2 (14%) and, finally, Vertisols with only 31 km2 (5%). The climatic index, known as evapotranspiration ratio (E/E0) is in adequate range for establishing systems with trees and grasses in cattle areas of the watershed. A list of tree species related to the prevailing soils is given. These should be considered for reforestation purposes with a view to improve the environmental and economic management of the region. Key words: silvopastoral systems, sustainable livestock.
Introducción
Al igual que en el resto de los países de América Latina, las diferentes prácticas utilizadas en Cuba para el desarrollo gana-dero a lo largo de los años, han desempe-ñado un papel determinante en el deterioro de los recursos naturales y dentro de ellas,
la deforestación ha sido sin dudas, una de las más perjudiciales (Acosta 2003).
Ante esta situación, el Estado cubano dedica especial interés a la reconversión de la ganadería, con la necesaria incorpo-ración de la dimensión ambiental en los planes de desarrollo ganadero, donde los

96 Zoe G. Acosta Gutiérrez
scientia-CUCBA 9(2):95—103.2007
sistemas silvopastoriles (SSP) han demos-trado constituir una alternativa promete-dora para el logro de una producción sos-tenible (Renda et al. 1997; Hernández et al. 1998; Instituto de Ciencia Animal 1998; Vargas 1998).
Por su importancia manifiesta, estos sistemas hoy forman parte de las acciones a desarrollar por el Ministerio de la Agricultura en Cuba a tenor de lo establecido en el Plan de Acción Nacional sobre la Diversidad Biológica (Ministerio de Ciencia, Tecnología y Medio Ambiente 2006).
������ ��� ��� ��� ��� ���
���
���
���
���
��� �� ����� �������������������������������������
����������
Figura 1. Mapa de la cuenca hidrográfica del San Pedro.
Objetivo
Contribuir a la extensión de SSP en las áreas ganaderas de la cuenca hidrográfica del río San Pedro en Camagüey, Cuba, me-diante la determinación de las áreas dispo-nibles por tipo de suelos y la propuesta de las especies que se pudieran emplear en la reforestación de las mismas.
Materiales y Métodos
La cuenca del río San Pedro se ubica en la vertiente sur de la provincia de Camagüey, Cuba; tiene una extensión de alrededor de 1052 km2 y abarca territorios de los muni-cipios Camagüey, Jimaguayú y Vertientes.

Sistemas silvopastoriles para una reconversión ambiental de la... 97
scientia-CUCBA 9(2):95—103.2007
Su río principal es el San Pedro el cual tiene una orientación suroeste-este, una longitud de 134 km y una pendiente media de 17% (figura 1).
Desde el punto de vista socioeconó-mico la cuenca reviste gran interés, por cuanto en la misma se localiza la ciudad de Camagüey (segunda del país por su exten-sión y tercera por el número de habitantes) y, además, se desarrolla una fuerte activi-dad ganadera, la cual representa el 63% del área total de la cuenca y está destinada al abasto de dicha ciudad así como a tributar grandes aportes al balance de alimentos para la Nación (figura 2).
En el presente estudio, se utilizó un SIG para determinar las áreas ganaderas que precisan ser reforestadas y para de-terminar las principales características físico-químicas de sus suelos. Para ello, se hicieron operaciones matemáticas y de Lenguaje Estructurado de Consulta (SQL) con los mapas base digital (1:50,000) y el de suelos (que incluye información sobre los tipos, la erosión, la humificación y la
profundidad efectiva de los mismos). Se tuvo en consideración la variación climáti-ca dentro de la cuenca.
A partir de la información derivada del análisis anterior, se confeccionó el lis-tado de las especies arbóreas maderable y frutales, promisorias para el desarrollo de SSP, de acuerdo con los principales trabajos publicados relativos a la regio-nalización de árboles y arbustos para la ganadería (Renda et al. op. cit.; Barreto et al. 1989; Hernández et al. op. cit.; Instituto de Ciencia Animal op. cit.; Oquendo 2002; Paretas 2002 y Paretas et al. 2002).
Resultados
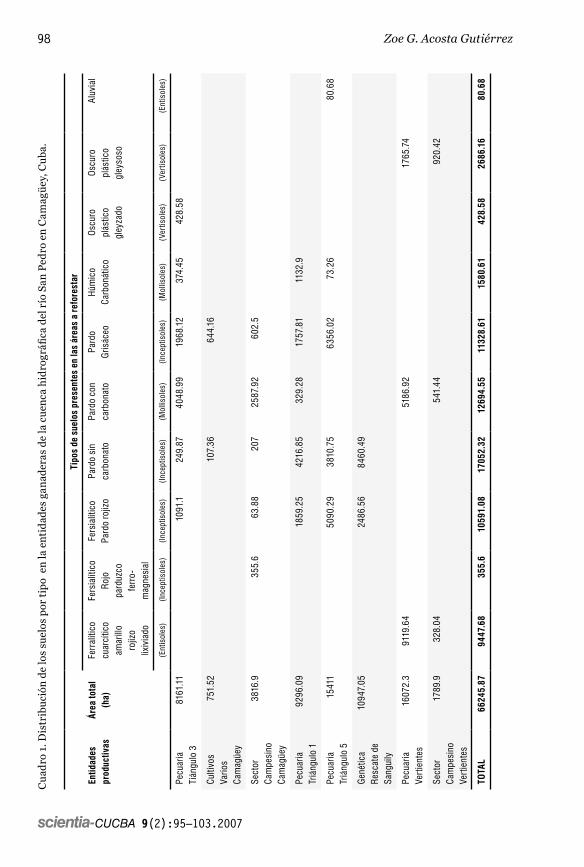
Las operaciones realizadas con las tablas contenidas en el SIG relacionadas con los suelos de la cuenca bajo estudio, arrojaron que la actividad ganadera cuenta con al-rededor de 662 km2 con posibilidades de ser reforestadas para un mejor desempeño ambiental y económico. El área se encuen-tra distribuida en ocho entidades produc-
� �� ����� �����������������������������������������������
���������������
������������������
������������������
Figura 2. Área de desarrollo ganadero en la cuenca del San Pedro.
98 Zoe G. Acosta Gutiérrez
scientia-CUCBA 9(2):95—103.2007
Cu
adro
1. D
istr
ibu
ción
de
los
suel
os p
or t
ipo
en
la e
nti
dad
es g
anad
eras
de
la c
uen
ca h
idro
gráfi
ca d
el r
ío S
an P
edro
en
Cam
agü
ey, C
ub
a.
Tipo
s de
sue
los
pres
ente
s en
las
área
s a
refo
rest
ar
Entid
ades
pr
oduc
tivas
Ár
ea to
tal
(ha)
Ferr
alíti
co
cuar
cític
o am
arill
o ro
jizo
lixiv
iado
Fers
ialít
ico
Rojo
pa
rduz
co
ferr
o-m
agne
sial
Fers
ialít
ico
Pard
o ro
jizo
Pard
o si
n ca
rbon
ato
Pard
o co
n ca
rbon
ato
Pard
o G
risác
eoH
úmic
o Ca
rbon
átic
oO
scur
o pl
ástic
o gl
eyza
do
Osc
uro
plás
tico
gley
soso
Aluv
ial
(Ent
isol
es)
(In
cept
isol
es)
(Inc
eptis
oles
)(I
ncep
tisol
es)
(Mol
lisol
es)
(Inc
eptis
oles
)(M
ollis
oles
)(V
ertis
oles
)(V
ertis
oles
)(E
ntis
oles
)
Pecu
aria
Ti
ángu
lo 3
8161
.11
1091
.124
9.87
4048
.99
1968
.12
374.
4542
8.58
Culti
vos
Vario
s Ca
mag
üey
751.
5210
7.36
644.
16
Sect
or
Cam
pesi
no
Cam
agüe
y
3816
.935
5.6
63.8
820
725
87.9
260
2.5
Pecu
aria
Tr
iáng
ulo
192
96.0
918
59.2
542
16.8
532
9.28
1757
.81
1132
.9
Pecu
aria
Tr
iáng
ulo
515
411
5090
.29
3810
.75
6356
.02
73.2
680
.68
Gen
étic
a Re
scat
e de
Sa
ngui
ly
1094
7.05
2486
.56
8460
.49
Pecu
aria
Ve
rtie
ntes
1607
2.3
9119
.64
5186
.92
1765
.74
Sect
or
Cam
pesi
no
Vert
ient
es
1789
.932
8.04
541.
4492
0.42
TOTA
L66
245.
8794
47.6
835
5.6
1059
1.08
1705
2.32
1269
4.55
1132
8.61
1580
.61
428.
5826
86.1
680
.68

Sistemas silvopastoriles para una reconversión ambiental de la... 99
scientia-CUCBA 9(2):95—103.2007
tivas en las que, a su vez, se desarrollan 40 estructuras de base entre Unidades Básicas de Producción Cooperativa (UBPC), Granjas Estatales y Cooperativas de Producción Agropecuaria (CPA).
El resumen acerca de la distribución de los suelos por tipo, se muestra en el cua-dro 1. Como se aprecia en el mismo, aque-llos que clasifican como Inceptisoles son predominantes en la región con 393 km2 para un 59% del área total a reforestar, seguidos por los Mollisoles con 143 km2 (22%), los Entisoles con 95 km2 (14%) y, por último, los Vertisoles con sólo 31 km2 (5%).
Por otra parte, la consulta al SIG arrojó que en las áreas donde se propone reforestar, la razón de evapotranspiración (E/E0) se comporta por encima de 0.5, lo cual según Riou (1992), resulta adecuado para el desarrollo de sistemas donde se combinen árboles, arbustos y pastos.
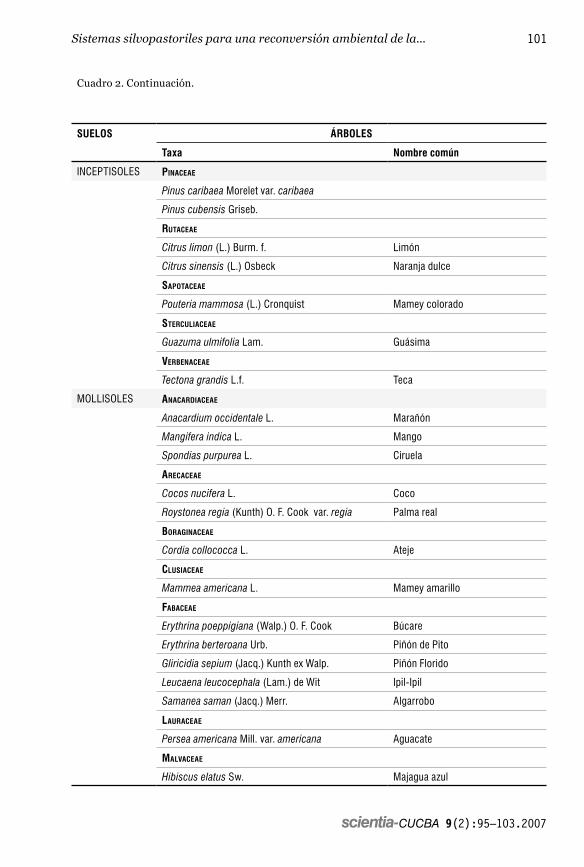
Las principales especies arbóreas tanto maderables como frutales que se pudieran establecer en las diferentes loca-lidades, por su afinidad o adaptación a los suelos, se listan en el cuadro 2. En la mis-ma se pone de manifiesto que hay variedad de especies reportadas en la literatura que son afines a los suelos presentes (Renda et al. op. cit.; Barreto et al. op. cit.; Hernández et al. op. cit.; Instituto de Ciencia Animal op. cit.; Oquendo op. cit.; Paretas op. cit. y Paretas et al. op. cit.), algunas de ellas, se encontraron además en la cuenca durante el diagnóstico de los recursos naturales de la misma (Primelles et al. 2004). Estas es-pecies se deberían considerar al momento de planificar la reforestación, lo que a su vez pudiera incrementar la diversidad de árboles en la región.
En tal sentido, el mayor número de especies arbóreas con posibilidades de establecimiento y desarrollo se localiza sobre suelos Mollisoles (29 especies) e Inceptisoles (27 especies) mientras que para los Entisoles y Vertisoles sólo se reco-mienda una especie por cada uno de ellos. En el cuadro 2 se presentan los nombres de las especies de acuerdo con la base de
datos w3TROPICOS en la red, enero 2008 (http://mobot.mobot.org/).
Conclusiones
• En la cuenca del río San Pedro la gana-dería dispone de un área significativa de suelos que precisan ser reforestados (alrededor de 662 km2), para contribuir a una mejor gestión ambiental y econó-mica de la región.
• Existe un grupo de especies arbóreas que por su afinidad con los suelos pre-sentes en la cuenca, se pudieran consi-derar para el establecimiento de siste-mas silvopastoriles.
Referencias
Acosta, Z. 2003. Los sistemas silvopastori-les: una práctica excelente para una ga-nadería sustentable. Módulo de Formación Ambiental Básica. Proyecto Acciones prio-ritarias para Consolidar la Protección de la Biodiversidad en el Ecosistema Sabana Camagüey. CUB/98/G32-CAPACIDAD 21. Pp. 5–6.
Barreto, A., C. Catases y Z. Acosta. 1989. Gramíneas y leguminosas naturales y/o naturalizadas de la provincia de Camagüey, Cuba. Rev. Pastos y Forrajes 21(1): 15–46.
Hernández, I., M. Milera, L. Simón, D. Hernández, J. Iglesias, L. Lamela, O. Toral, C. Matías y G. Francisco. 1998. Avances de las investigaciones en siste-mas silvopastoriles en Cuba. I Conferencia Electrónica sobre Agroforestería para la Producción Animal en América Latina (Agrofor1). (http://www.cipav.org.co/agro-for).
Instituto De Ciencia Animal. Grupo Multidisciplinario de Leguminosas. 1998. Enfoque acerca del trabajo sobre ár-boles y arbustos desarrollados por el institu-to de ciencia animal de Cuba. I Conferencia Electrónica sobre Agroforestería para la Producción Animal en América Latina (Agrofor1). (http://www.cipav.org.co/agro-for).
Ministerio de Ciencia, Tecnología y Medio Ambiente. 2006. Plan de Acción Nacional 2006/2010 sobre la Diversidad Biológica. Centro Nacional de Seguridad Biológica, La Habana, Cuba 64 pp.

100 Zoe G. Acosta Gutiérrez
scientia-CUCBA 9(2):95—103.2007
Cuadro 2. Listado de posibles relaciones especies-suelo para el establecimiento de nuevos SSP en la cuenca hidrográfica del San Pedro.
SUELOS ÁRBOLES
Taxa Nombre común
ENTISOLES ARECACEAE
Coccothrinax miraguama (Kunth) León ssp.
miraguama
Miraguano
INCEPTISOLES ANACARDIACEAE
Anacardium occidentale L. Marañón
Mangifera indica L. Mango
Spondias purpurea L. Ciruela
Cocos nucifera L. Coco
Roystonea regia (Kunth) O. F. Cook var. regia Palma real
BORAGINACEAE
Cordia collococca L. Ateje
Cordia gerascanthus L. Baría
CLUSIACEAE
Mammea americana L. Mamey amarillo
FABACEAE
Erythrina poeppigiana (Walp.) O. F.Cook Búcare
Erythrina berteroana Urb. Piñón de Pito
Gliricidia sepium (Jacq.) Kunth ex Walp. Piñón Florido
Leucaena leucocephala (Lam.) de Wit Ipil-Ipil
Samanea saman (Jacq.) Merr. Algarrobo
Tamarindus indica L. Tamarindo
LAURACEAE
Persea americana Mill. var. americana Aguacate
MELIACEAE
Cedrela odorata L. Cedro
Swietenia macrophylla King Caoba de Honduras
Swietenia mahagoni (L.) Jacq.* Caoba Antillana
Trichilia hirta L. Cabo de hacha
MYRTACEAE
Eucalyptus saligna Sm. Eucalipto
* Checkl. Sp. [2003]. CITES Appendix II - Comercio controlado para evitar un uso incompa-tible con la sobrevivencia de la especie – Global Conservation System. UNEP World Conservation Monitoring Centre, Cambridge.
Sistemas silvopastoriles para una reconversión ambiental de la... 101
scientia-CUCBA 9(2):95—103.2007
Cuadro 2. Continuación.
SUELOS ÁRBOLES
Taxa Nombre común
INCEPTISOLES PINACEAE
Pinus caribaea Morelet var. caribaea
Pinus cubensis Griseb.
RUTACEAE
Citrus limon (L.) Burm. f. Limón
Citrus sinensis (L.) Osbeck Naranja dulce
SAPOTACEAE
Pouteria mammosa (L.) Cronquist Mamey colorado
STERCULIACEAE
Guazuma ulmifolia Lam. Guásima
VERBENACEAE
Tectona grandis L.f. Teca
MOLLISOLES ANACARDIACEAE
Anacardium occidentale L. Marañón
Mangifera indica L. Mango
Spondias purpurea L. Ciruela
ARECACEAE
Cocos nucifera L. Coco
Roystonea regia (Kunth) O. F. Cook var. regia Palma real
BORAGINACEAE
Cordia collococca L. Ateje
CLUSIACEAE
Mammea americana L. Mamey amarillo
FABACEAE
Erythrina poeppigiana (Walp.) O. F. Cook Búcare
Erythrina berteroana Urb. Piñón de Pito
Gliricidia sepium (Jacq.) Kunth ex Walp. Piñón Florido
Leucaena leucocephala (Lam.) de Wit Ipil-Ipil
Samanea saman (Jacq.) Merr. Algarrobo
LAURACEAE
Persea americana Mill. var. americana Aguacate
MALVACEAE
Hibiscus elatus Sw. Majagua azul

102 Zoe G. Acosta Gutiérrez
scientia-CUCBA 9(2):95—103.2007
Cuadro 2. Continuación.
SUELOS ÁRBOLES
Taxa Nombre común
MOLLISOLES MELIACEAE
Cedrela odorata L.
Swietenia macrophylla King
Swietenia mahagoni (L.) Jacq.*
Trichilia hirta L.
MYRTACEAE
Eucalyptus saligna Sm. Eucaliptu
Pisidium guajava L. Guayaba
PINACEAE
Pinus caribaea Morelet var. caribaea Pino
Pinus cubensis Griseb. Pino
RUTACEAE
Citrus limon (L.) Burm. F. Limón
Citrus sinensis (L.) Osbeck Naranja dulce
SAPOTACEAE
Manilkara sapota (L.) van Royen Níspero
Pouteria mammosa (L.) Cronquist Mamey colorado
STERCULIACEAE
Guazuma ulmifolia Lam. Guásima
VERBENACEAE
Tectona grandis L. Teca
VERTISOLES STERCULIACEAE
Guazuma ulmifolia Lam. Guásima
* Checkl. Sp. [2003]. CITES Appendix II - Comercio controlado para evitar un uso incompatible con la
sobrevivencia de la especie – Global Conservation System. UNEP World Conservation Monitoring Centre,
Cambridge.

Sistemas silvopastoriles para una reconversión ambiental de la... 103
scientia-CUCBA 9(2):95—103.2007
Oquendo, G. 2002. Tecnologías para el fo-mento y explotación de pastos y forrajes. Eds. Agro Acción Alemana, Ministerio Federal para la Cooperación Económica y el Desarrollo y Asociación Cubana de Producción Animal, La Habana, Cuba, 112 pp.
Paretas, J. J. 2002. Sostenibilidad ganadería-medio ambiente basada en PAF. ACPA 4: 22–23.
Paretas, J. J., M. López, R. Acosta, R. Serrano y L. Gallardo. 2002. Regionalización de árboles multipropósi-tos. En: Memoria del V Taller Internacional Silvopastoril y I Reunión Regional de Morera, Matanzas, Cuba.
Primelles, J., M.E. Zequeira, Z. Acosta, G. Martín, A. Ferrer, O. Brito, L. Hernández, M.E. Li, D. Francis, A. Líaz, W. Machado, J. Aguilar, J.M. Plasencia, D. Godínez, N. Enríquez, A. Cebrián y X. López. 2004. Cuenca del Río San Pedro: Diagnóstico ambiental y
objetivos para su manejo integrado. Revista Medio Ambiente y Desarrollo. AMA. CUBA. Año 4, No. 7. ISSN: 1683-8904.
Renda, A., E. Calzadilla, M. Jiménez y J. Sánchez. 1977. La Agroforestería en Cuba. Red Latinoamericana de Cooperación Técnica en Sistemas Agroforestales. Dirección de Recursos Forestales, FAO, Roma. Oficina Regional de la FAO para América Latina y el Caribe, Santiago de Chile, 63 pp.
Riou, C. 1993. Case study on instrument for the diagnosis of dryness: frequential análisis of water déficit. Franquin model In: Monitoring assessment and combat of dranght and de-sertification. Report, Geneva (47): 12–24.
Vargas, S. 1998. Los sistemas agroecológicos en la producción ganadera. En Producción bovina sostenible. Ed. ACPA, La Habana, Cuba, 135 pp.
FECHA EFECTIVA DE PUBLICACIÓN 29 DE DICIEMBRE DE 2007

104
scientia-CUCBA 9(2):104.2007

scientia-CUCBA 9(2):105—117.2007
Extracción de ADN en accesiones de maíz y teocintle
Martha Isabel Torres-Morán, Moisés Martín Morales-Rivera, José de Jesús Sánchez-González, Lino De la Cruz-Larios, José Ron-Parra y Fernando Santacruz-RuvalcabaInstituto de Manejo y Aprovechamiento de Recursos Fitogenéticos. Departamento de Producción Agrícola. Centro Universitario de Ciencias Biológicas y Agropecuarias. Universidad de Guadalajara. Km 15.5 Carretera a Nogales. C.P. 45110 Las Agujas, Zapopan, Jalisco. Correo electrónico: [email protected]
Resumen: Con la finalidad de estudiar métodos de extracción de ADN y definir un protocolo para accesiones de teocintle, se probaron tres métodos de extracción en 28 accesiones, incluyendo dos líneas homocigóticas de maíz y la cruza simple entre ellas. Se evaluó la calidad y cantidad del ADN obtenido por cada método utilizando espectrofotometría y electroforesis, además se realizó una caracterización espectrofotométrica de las muestras. No se detectaron diferencias significativas en cuanto a concentración y calidad de ADN entre teocintle y maíz ni efectos diferenciales debido a los métodos, lo cual indica que los resultados de esta investigación se pueden usar de manera amplia en el resto de especies y poblaciones del género Zea. Los diferentes métodos empleados para la evaluación de la concentración y calidad del ADN obtenido, fueron consistentes y se complementan como auxiliares en la elección de los protocolos apropiados para la extracción de ADN en el género Zea. Se probó además la amplificación del ADN extractado utilizando un cebador aleatorio de RAPD, el cual detectó polimorfismo en las diferentes accesiones.
Palabras clave: Extracción ADN, caracterización espectrofotométrica, teocintle.
Abstract: In order to study DNA extraction methods and to define an appropriate protocol for isolating DNA in teosinte, three isolation methods were tested in twenty-eight accessions, including two homozygotic lines and their single cross. The quality and quantity of DNA in samples were evaluated for each method using spectrophotometric and electrophoretic techniques. In addition, spectrophotometrical characterization was made with samples obtained. Based on the above criteria, no significant differences in quality and quantity among accessions and no differential effects due to the methods were found. Thus, the results yielded in the research can be amply used for the remaining species in this genus and in populations of the Zea genus. All methods used for the evaluation of quality and quantity of DNA obtained[including spectrofotometrical,] were consistent, and the results can be used as a complementary tool for choosing an adequate method for isolating DNA in this genus. Moreover, the amplification pattern was tested by means of RAPD markers, resulting in the detection of polymorphic bands among accessions.
Key words: DNA isolation, spectrophotometrical characterization, teosinte.
Introducción
El avance de las ciencias genómicas en los últimos años, constituye una herramien-ta valiosa que incide en los campos de la conservación y aprovechamiento de los recursos fitogenéticos. En la actualidad se encuentran disponibles técnicas que
permiten analizar diferencias en el ámbito molecular en cualquier organismo y han demostrado su utilidad en la caracteriza-ción e identificación de partes especificas del genoma.

106 Martha Isabel Torres-Morán et al.
scientia-CUCBA 9(2):105—117.2007
Con el desarrollo de la Reacción en Cadena de la Polimerasa (PCR), surgieron numerosas técnicas, llamadas en forma genérica Marcadores Moleculares, que permiten la caracterización para el mane-jo, aprovechamiento y evaluación de los recursos fitogenéticos.
La obtención de ADN de buena ca-lidad, es el primer paso para la aplicación de técnicas moleculares que persigan fines específicos, tales como las estimaciones de la diversidad y estructura genética de po-blaciones o relaciones filogenéticas entre especies.
La importancia de elegir métodos apropiados de extracción de ADN, reside no solo en la cantidad que se obtenga, sino en su pureza. Algunas especies contienen altos niveles de polisacáridos y metabolitos secundarios que pueden inhibir el fun-cionamiento de enzimas de restricción y complicar el buen funcionamiento de mé-todos basados en PCR tales como RAPDs (Random Amplifield Polymorphic DNA), AFLPs (Amplifield Fragment Length Polymorphism) y otras técnicas molecula-res (Csaikl et al. 1998).
Hoy día se han desarrollado bastan-tes métodos para aislar ADN de alto peso molecular a partir de diferentes tejidos de plantas. Ellos difieren en el tejido que uti-lizan para la extracción, el cual puede ser fresco, liofilizado, congelado y en cantida-des que dependen de la condición del tejido (Hillis et al. 1996). También pueden diferir en el tipo de compuestos utilizados para la lisis, en la precipitación de las proteínas y en la purificación del ADN; en muchos casos ha sido necesario probar sustancias que ayuden a eliminar las contaminaciones con polisacáridos y compuestos fenólicos que varían según la especie de que se trate (Valadez y Kahl 2000; Csaikl et al. 1998; Drábková et al. 2002; Rogstad 2003).
Por último, el protocolo elegido, debe garantizar eficiencia, reproducibilidad y estabilidad del ADN, así como potencial para aplicar el producto en amplificacio-nes, restricciones, formación de bancos genómicos, estudios filogenéticos, entre
otras (Wolfe et al. 2002). Se sabe que la extracción de ADN es muy difícil, sobre todo en algunas especies forestales (Csaikl et al. 1998). En maíz no se han reportado dificultades con los métodos de extracción más comunes, sin embargo, tampoco se conocen hasta el momento informes sobre métodos específicos de extracción de ADN para las diferentes especies silvestres del género Zea, relacionadas al maíz.
Por tanto, el objetivo de la presente investigación fue evaluar la calidad y can-tidad de ADN obtenido de accesiones de teocintle y de maíz, con base en tres méto-dos de extracción, y verificar la amplifica-ción por PCR de fragmentos obtenidos con RAPD.
Materiales y métodos
El material genético que se usó consistió de 28 accesiones representantes de la mayoría de las especies conocidas del género Zea, las cuales se listan en el cuadro 1. Los tra-bajos de laboratorio se llevaron a cabo en dos etapas; en la primera se utilizaron cin-co accesiones de tres especies de teocintle y dos líneas homocigóticas de maíz, así como la cruza simple entre dichas líneas, ya que en maíz se tienen reportes de métodos de extracción eficientes en que se obtuvieron buenas cantidades de ADN (cuadro 1). En la segunda etapa se usaron 20 materiales que incluyeron 15 accesiones de teocint-le y cinco líneas homocigóticas de maíz. Los materiales fueron sembrados durante el ciclo de verano del 2005 en el Campo Experimental del Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA) de la Universidad de Guadalajara, en Nextipac, Zapopan, Jalisco.
Se tomaron muestras frescas de cada uno de los materiales indicados en el cua-dro 1, consistentes en una sección de alre-dedor de 12 cm de la parte media de hojas sanas de plantas de alrededor de 30 días de germinadas las semillas.
En la etapa 1, se utilizaron los méto-dos de extracción reportados por Clarke et al. (1989), Saghai-Maroof et al. (1984)

Extracción de ADN en accesiones de maíz y teocintle 107
scientia-CUCBA 9(2):105—117.2007
Cuadro 1. Accesiones del género Zea utilizadas en la extracción de ADN.
No. Accesión Especie Raza/material Procedencia
Etapa 1
1 LUG-14 Zea mays L. ssp. mays Línea Proyecto Maíz CUCBA
2 LUG-03 Zea mays ssp. mays Línea Proyecto Maíz CUCBA
3 JSG-RMM-LCL-480 Zea mays ssp. mexicana
(Schard)H.H. Iltis
Chalco Cocotitlán, México
4 JSG-RMM-LCL-450 Zea mays ssp. mexicana Mesa Central San Agustín del Maíz,
Michoacán
5 JSG-RMM-LCL-500 Zea mays ssp. parviglumis
H.H. Iltis & Doebley
Balsas Zacatlancillo, Guerrero
6 JSG-RMM-LCL-555 Zea mays ssp. parviglumis Balsas Talpitita
7 JSG-RMM-LCL-551 Zea diploperennis H.H. Iltis,
Doebley & R. Guzmán
Z. diploperennis Las Joyas, Jalisco
8 LUG-14 x LUG-03 Zea mays ssp. mays Híbrido Proyecto Maíz CUCBA
Etapa 2
9 NC-354 Zea mays ssp. mays Línea North Carolina State U.
10 LUG-14 Zea mays ssp. mays Línea Proyecto Maíz CUCBA
11 LUG-03 Zea mays ssp. mays Línea Proyecto Maíz CUCBA
12 LUG-282 Zea mays ssp. mays Línea Proyecto Maíz CUCBA
13 W22 Zea mays ssp. mays Línea University of Wisconsin
14 JSG-RMM-LCL-495 Zea mays ssp. parviglumis Balsas Ixcateopan, Gro.
15 JSG Y ERV-409 Zea mays ssp. mexicana Mesa Central Ahuatlán-Poncitlán, Jal.
16 JSG Y RMM-441 Zea mays ssp. mexicana Mesa Central Puruándiro, Mich.
17 JSG Y RMM-450 Zea mays ssp. mexicana Mesa Central San Agustín del Maíz,
Michoacán
18 JSG Y RMM-451 Zea mays ssp. mexicana Mesa Central Chucándiro, Mich.
19 JSG-RMM-LCL-536 Zea mays ssp. mexicana Mesa Central Cd. Hidalgo, Mich.
20 JSG Y RMM-463 Zea mays ssp. mexicana Chalco Opopeo, Mich.
21 JSG-433 Zea mays ssp. mexicana Chalco Aljojuca, Puebla
22 JSG-RMM-LCL-471 Zea mays ssp. mexicana Chalco Chapultepec, México
23 JSG-RMM-LCL-477 Zea mays ssp. mexicana Chalco San Pedro Nexapa, Méx.
24 JSG-RMM-LCL-479 Zea mays ssp. mexicana Chalco Amecameca, Méx.
25 JSG Y LOS-75 Zea mays ssp. mexicana Chalco Cojumatlán, Mich.
26 JSG-RMM-LCL-552 Zea diploperennis Z. diploperennis Corralitos, Jal.
27 Agua Blanca Zea luxurians (Durieu & Asch.)
R.M. Bird
Guatemala Agua Blanca, Guatemala
28 Monajil Z. mays ssp. huehuetenansis
(H.H. Iltis & Doebley) Doebley
Huehuetenango Monajil, Guatemala
LUG = Línea Universidad de Guadalajara

108 Martha Isabel Torres-Morán et al.
scientia-CUCBA 9(2):105—117.2007
y Keb-Llanes et al. (2002); en el cuadro 2 se citan las diferencias entre los métodos. Cada muestra se procesó por los tres mé-todos y se hicieron tres repeticiones con cada de ellos. En la etapa 2, el método de extracción usado fue el de Saghai-Maroof et al. (op. cit.)
Para la evaluación de calidad y can-tidad del ADN, se realizaron lecturas de absorbancia de las muestras con un espec-trofotómetro Jenway 6305 UV/VIS a una longitud de onda de 260 nm. Se determinó la concentración con la fórmula:
ConcADN(µg/µl)=[DO260 × fd × 50µg/ml]/1000
donde el factor de dilución ( fd) fue 1:200 y el término 50 µg/ml es debido a que una densidad óptica a 260 nm (DO260)=1 corresponde a una concentración de 50 microgramos (µg) por microlitro (µl) de ADN. Por otra parte, con lecturas en 280 nm puede estimarse la cantidad de com-puestos fenólicos y proteínas. La relación
entre esas dos lecturas (DO260/280) aporta una estimación de la pureza del ácido nu-cléico obtenido; las preparaciones puras, muestran valores de entre 1.8 y 2.0 en esta relación (Valadez y Kahl 2000). Con el fin de tener una caracterización espectrofoto-métrica detallada, se realizaron lecturas de 230 a 300 nm, en donde la lectura de 240 nm indica cantidad de carbohidratos presentes en la misma.
Las variables medidas fueron: densi-dad óptica de las muestras a 260 y 280 nm de donde posteriormente se infiere concen-tración de ADN y presencia de compuestos fenólicos contaminantes respectivamente. Se estimó la relación DO260/280 como indi-cador de calidad, según lo reportado por Csaikl, et al. (1998).
Se verificó la calidad del ADN extraí-do, al utilizar todas las muestras y repeti-ciones, en un gel de agarosa 1%.
Se llevó a cabo una amplificación del ADN extractado por los métodos de Saghai-Maroof y Keb-Llanes, mediante
Cuadro 2. Comparación de los pasos utilizados en cada método de extracción de ADN.
Pasos Keb-Llanes et al., (2002)
Saghai-Maroof et al.,(1984)
Clarke et al., (1989)
Lisis celular
Triturado de las muestras N2 N2 N2
Número de Buffer de extracción utilizados 2 1 1
Precipitación de proteínas
Temperatura y tiempo en baño María 65° C por 10’ 60°C por 30’ 65°C por 40’
Eliminación de residuos de tejido centrifugación centrifugación centrifugación
Precipitación de ADN
Precipitación ADN isopropanol isopropanol Etanol 100%
Lavado y resuspensión de ADN
Lavados Etanol 70% Etanol de 3-4 Etanol al 70% 3
Resuspensión del ADN 100µL T.E. 50-100µL T.E. 200µL T.E
Tiempo total aproximado para 8 muestras 6 hrs 4 hrs. 5hrs

Extracción de ADN en accesiones de maíz y teocintle 109
scientia-CUCBA 9(2):105—117.2007
un kit para RAPD Amersham, y el cebador 5’d[GTTTCGCTCC’]3’ en las condiciones reportadas en el kit Ready-to-go RAPD Análisis Beads® (Amersham 2003).
DISEÑO EXPERIMENTAL
Para la etapa 1, el diseño experimental fue de bloques al azar con arreglo factorial de los tratamientos. Los factores evaluados fueron los métodos de extracción (Clarke, Saghai-Maroof y Keb-Llanes) y ocho ac-cesiones del género Zea. Con los datos obtenidos de las lecturas 260 y 280 nm se efectuó un análisis de varianza. En los casos de falta de interacción, se determina-ron las diferencias significativas entre los métodos o entre las accesiones con base en la prueba de Tukey (P ≤0.05). Para la etapa 2, el diseño experimental fue de bloques al azar con 20 accesiones y siete plantas por accesión.
CRITERIOS PARA LA ELECCIÓN DEL MÉTODO
Existen factores de gran importancia que influyen en la elección del método y que fueron tomados en cuenta en el presente trabajo, son:• La cantidad de ADN obtenido, que para
condiciones de almacenamiento debe ser del orden de 700 ng/μl en adelante (Valadez y Kahl op. cit.).
• La calidad de ADN, indicada por menor presencia de contaminantes fenólicos, polisacáridos y ARN que inhiben la ac-ción de enzimas de restricción y polime-rasas en reacciones posteriores de am-
plificación (Murray et al. 1996; Csaikl et al. op. cit.; Rogstad 2003).
• Facilidad y costos. Estos factores de-penden en gran medida del número de pasos que se realicen en la extracción. Algunos métodos utilizan varios pasos de precipitación de proteínas, como es el caso del método Keb-Llanes, et al. (op. cit.), o varios pasos de lavado y precipi-tación del ADN. Los costos dependen de la cantidad de reactivos empleados y el tiempo que se utiliza en el procesamien-to de las muestras.
• Reproducibilidad. El método debe ga-rantizar la obtención de cantidad y cali-dad constante en las muestras procesa-das, así como la estabilidad del ADN en almacenamiento (Csaikl et al. op. cit.).
Resultados y discusión
Se realizó la electroforesis en gel de aga-rosa 1%, utilizando las muestras y repe-ticiones de la etapa 1, obteniéndose las bandas características de la presencia de ADN de alto peso molecular en todos los casos (figura 1) la presencia de barrido en las muestras, indica contenido probable de polisacáridos y residuos fenólicos. Existen estudios sobre la inhibición que pueden ejercer los polisacáridos en los análisis RAPD (Pandey y Flournoy 1996) por lo cual es importante la verificación de la amplificación del ADN obtenido, al utilizar dicho marcador molecular.
En el cuadro 3 se muestran las lec-turas de absorbancia a las longitudes de onda de 260 y 280 nanómetros, así como
Figura 1. Electroforesis en gel de agarosa 1% correspon-dientes a ADN de alto peso molecular para 5 accesiones de Teocintle, dos líneas homo-cigóticas de maíz y su cruza siemple. Marcador de peso molecular 22 kb (M).

110 Martha Isabel Torres-Morán et al.
scientia-CUCBA 9(2):105—117.2007
la relación entre esas dos lecturas como indicador de calidad del ADN.
ANÁLISIS DE VARIANZA ETAPA 1
Con los datos de calidad de ADN (DO a 280nm), relación DO260/280 y concen-tración de ADN se realizó un análisis de varianza, cuyos resultados se presentan en el cuadro 4. El análisis de varianza de-tectó diferencias altamente significativas (P<0.01) entre repeticiones para las tres variables consideradas. Por su parte, se de-tectaron diferencias significativas (P<0.05) para la calidad de ADN (DO a 280nm) y para la concentración de ADN entre los métodos (M), sin embargo, no se detec-taron diferencias entre las accesiones (A) estudiadas ni para la interacción M × A. Para la relación DO260/280 no se detectaron
diferencias significativas entre métodos, ni entre accesiones o la interacción.
La comparación de medias de los mé-todos de extracción se muestra en el cuadro 5. La comparación estadística indica que el método con el cual se obtuvo una mayor concentración de ADN es el método de Saghai-Maroof et al. (op. cit.), con el cual se obtuvo en promedio 2085 μg/mL. Aún cuando no se detectaron diferencias signi-ficativas entre las accesiones estudiadas, cabe señalar que la cruza simple LUG-03 × LUG-14 fue el material del que se obtuvo la mayor cantidad de ADN con un promedio de 2275 μg/mL.
De manera similar que para con-centración de ADN, hubo diferencias alta-mente significativas entre los métodos de extracción; sin embargo, no se detectaron diferencias entre las accesiones estudiadas
Cuadro 3. Datos generales de concentración de ADN (µg/mL) y relación DO260/280 del género Zea por tres métodos de extracción.
Accesiones de acuerdo al cuadro 1
Métodos Rep. 1 2 3 4 5 6 7 8
Clarke I Conc ADN 1130 1420 1690 1520 2200 2080 1600 1440
DO260/280 1.5 1.6 1.6 1.6 1.6 1.6 1.6 1.7
II Conc ADN 1240 1550 1300 1790 1490 1880 1540 1510
DO260/280 1.7 1.7 1.6 1.7 1.7 1.7 1.6 1.7
III Conc ADN 1150 2510 960 1360 1640 780 1110 4780
DO260/280 1.4 1.7 1.7 1.6 1.3 1.6 1.7 1.7
Keb-Llanes I Conc ADN 870 1710 700 1080 1340 810 1320 1730
DO260/280 1.8 1.8 1.8 1.8 1.7 1.7 1.6 1.7
II Conc ADN 800 1360 950 1160 1220 950 1570 1240
DO260/280 1.6 1.6 1.6 1.8 1.7 1.6 1.7 1.8
III Conc ADN 560 1270 2680 1610 1000 1640 1920 1130
DO260/280 1.8 1.7 1.7 1.7 1.5 1.6 1.4 1.7
Saghai-Maroof I Conc ADN 1930 780 780 2900 480 1930 480 2540
DO260/280 1.7 1.7 1.6 1.7 1.8 1.6 1.8 1.7
II Conc ADN 580 1140 1090 1190 1090 1140 1910 1430
DO260/280 1.5 1.6 1.5 1.6 1.5 1.6 1.7 1.6
III Conc ADN 2020 2860 2730 2760 2770 3970 4080 3370
DO260/280 1.8 1.7 1.7 1.7 1.6 1.7 1.5 1.6

Extracción de ADN en accesiones de maíz y teocintle 111
scientia-CUCBA 9(2):105—117.2007
ni para la interacción. El método con el que se obtuvo significativamente la mejor calidad de ADN (cuadro 5), es decir sin presencia de ARN, fenoles y proteínas, es el método de Keb-Llanes et al. (op. cit.), con un promedio 0.171. El promedio de lectura más bajo de DO a 280 nm indica menor presencia de contaminantes y por lo tanto ADN más puro. La muestra de donde se obtuvo el ADN con mejor calidad perte-neció a LUG 14 (0.154)
En este punto es de importancia re-cordar que no se detectaron diferencias significativas entre accesiones ni se detectó interacción entre los métodos y las accesio-nes, para la concentración de ADN ni para la calidad del ADN. Dichos resultados son de gran importancia dado que indican que los métodos se podrán elegir de manera general sin esperar diferencias entre o dentro de las especies de Zea. La diferen-cia en calidad y cantidad de ADN obtenido en los métodos, indica que a pesar de que se encuentren disponibles varios protoco-los de extracción, la elección de cualquiera de ellos, dependerá de los factores involu-crados en el análisis, como costo, tiempo invertido en la extracción y facilidad de aplicación del método. La calidad del ADN es importante, en algunas técnicas mo-leculares que requieren de muestras más puras, como es el caso de los marcadores basados en hibridación “Southern Blot”.
De acuerdo con Lynne (1998) la ca-lidad del ADN extraído puede afectar de manera directa el resultado que se obtenga
en los experimentos posteriores, debido a la presencia de nucleasas contaminantes y degradación que pueden inhibir la ma-nipulación biológica subsecuente. Esta misma autora menciona que extraer ácidos nucleicos con la calidad requerida, no es un asunto trivial y las técnicas deben ser optimizadas para cada caso particular.
ANÁLISIS DE VARIANZA ETAPA 2
El análisis de varianza para la calidad de ADN (DO a 280nm) y para la concentra-ción de ADN se presenta en el cuadro 6. El análisis de varianza detectó diferencias altamente significativas (P<0.01) única-mente entre plantas para las dos variables consideradas. Estos resultados confirman lo encontrado en el experimento de la etapa 1, es decir el método de extracción empleado es similar en cuanto a la cali-dad y concentración del ADN extraído, sin importar las poblaciones o especies dentro del género Zea. Aún cuando no hay diferencias significativas entre las accesio-nes estudiadas, hay algunas poblaciones de teocintle que presentaron tendencias de mejor concentración del ADN extraído (cuadro 7); tal es el caso de los teocintles de Poncitlán, Jalisco; Ixcateopan, Guerrero y Amecameca, Estado de México. Respecto a calidad del ADN, los menores valores de DO 280, se presentaron en las líneas NC-354, W22 y el teocintle de Puruándiro, Michoacán.
Cuadro 4. Análisis de varianza para calidad de ADN (DO a 280nm), relación DO260/280 y concen-tración de ADN en ocho accesiones del género Zea por tres métodos de extracción.
DO 280 DO 260/280 Concentración ADN
F.V. G.L. C.M. Pr >F C.M. Pr >F C.M. Pr >F
Repetición 2 0.0709 <.0001 0.2133 <.0001 7344866.7 <.0001
Método (M) 7 0.0222 0.031 0.0147 0.166 1995116.7 0.024
Accesión (A) 2 0.0066 0.368 0.0063 0.587 596562.5 0.318
M x A 14 0.0064 0.402 0.0062 0.678 547719.1 0.376
Error 46 0.0059 0.0079 494193.8
CV (%) 38.7 5.0 39.4

112 Martha Isabel Torres-Morán et al.
scientia-CUCBA 9(2):105—117.2007
Cuadro 5. Medias de accesiones y métodos de extracción para calidad de ADN (DO a 280 nm), relación DO260/280 y concentración de ADN para 8 accesiones del género Zea.
Accesión Clarke Keb-Llanes Shagai-Maroof Media
DO 280
JSG-RMM-LCL-450 0.225 0.133 0.265 0.208
JSG-RMM-LCL-480 0.217 0.129 0.190 0.179
JSG-RMM-LCL-500 0.203 0.190 0.206 0.199
JSG-RMM-LCL-551 0.183 0.156 0.267 0.202
JSG-RMM-LCL-555 0.230 0.109 0.259 0.199
LUG-03 0.171 0.218 0.207 0.199
LUG-14 0.154 0.130 0.178 0.154
LUG-14xLUG-03 0.171 0.300 0.278 0.250
Media 0.194ab 0.171b 0.231a
DO 260/ 280
JSG-RMM-LCL-450 1.812 1.772 1.858 1.814
JSG-RMM-LCL-480 1.806 1.736 1.803 1.782
JSG-RMM-LCL-500 1.800 1.753 1.777 1.777
JSG-RMM-LCL-551 1.759 1.765 1.708 1.744
JSG-RMM-LCL-555 1.802 1.770 1.805 1.792
LUG-03 1.821 1.757 1.663 1.747
LUG-14 1.791 1.665 1.831 1.762
LUG-14xLUG-03 1.816 1.799 1.818 1.811
Media 1.801 1.752 1.783
Concentración de ADN
JSG-RMM-LCL-450 2053 1172 2437 1887
JSG-RMM-LCL-480 1972 1117 1720 1603
JSG-RMM-LCL-500 1817 1680 1867 1788
JSG-RMM-LCL-551 1607 1373 2368 1783
JSG-RMM-LCL-555 2070 955 2337 1787
LUG-03 1557 1940 1823 1773
LUG-14 1378 1087 1617 1361
LUG-14xLUG-03 1557 2760 2508 2275
Media 1751ab 1510b 2085a
* Medias con la misma letra por renglón son estadísticamente iguales entre sí (Tukey, 0.05).

Extracción de ADN en accesiones de maíz y teocintle 113
scientia-CUCBA 9(2):105—117.2007
Cuadro 6. Análisis de varianza para calidad de ADN (DO a 280nm, y concentración de ADN en 20 accesiones del género Zea con base en el método de Saghai-Maroof et al. 1984.
DO 280 Concentración
ADN
F.V. G.L. C.M. Pr > F C.M. Pr > F
Planta 6 0.110077 0.001 8608212.4 <.0001
Accesión 19 0.033736 0.245 1128651.3 0.800
Error 114 0.027395 1585029.5
Total 139
CV(%) 39.2 35.0
CARACTERIZACIÓN ESPECTROFOTOMÉTRICA DE LAS MUESTRAS
La caracterización espectrofotométrica de las muestras de ADN extraído, es un procedimiento que permite ver en forma gráfica la calidad y cantidad que se tiene en las muestras analizadas. Una curva tí-pica de ADN con buena calidad y cantidad, es aquella que muestra valores de DO ba-jos para las lecturas 240 y 280 nm y altos para la lectura 260 nm. Los resultados de la caracterización espectrofotométrica se presentan en las figuras 1 a 3, para las ac-cesiones 1 a 8 del cuadro 1.
Los resultados de la caracterización con el método Clarke et al. (op. cit.), se presenta en la figura 1. Cabe señalar que
Cuadro 7. Medias de 20 accesiones para calidad de ADN (DO a 280nm) y concentración de ADN con base en el método de extracción de Saghai-Maroof et al., 1984.
Accesión Origen Especie DO280 Conc.ADN
NC-354 North carolina State Univ. Z. mays ssp. mays 0.233 3716
LUG-14 Proyecto Maíz CUCBA Z. mays ssp. mays 0.433 3741
LUG-03 Proyecto Maíz CUCBA Z. mays ssp. mays 0.415 3631
LUG-282 Proyecto Maíz CUCBA Z. mays ssp. mays 0.419 3729
W22 University of Wisconsin Z. mays ssp. mays 0.301 2924
JSG-RMM-LCL-495 Ixcateopan, Gro. Z. mays ssp. parviglumis 0.497 4324
JSG Y ERV-409 Ahuatlán-Poncitlán, Jal. Z. mays ssp. mexicana 0.521 4554
JSG Y RMM-441 Puruándiro, Mich. Z. mays ssp. mexicana 0.350 3033
JSG Y RMM-450 S Agustín del Maíz, Mich. Z. mays ssp. mexicana 0.431 3660
JSG Y RMM-451 Chucándiro, Mich. Z. mays ssp. mexicana 0.404 3302
JSG-RMM-LCL-536 Cd. Hidalgo, Mich. Z. mays ssp. mexicana 0.413 3451
JSG Y RMM-463 Opopeo, Mich. Z. mays ssp. mexicana 0.496 3437
JSG-433 Aljojuca, Pue. Z. mays ssp. mexicana 0.412 3271
JSG-RMM-LCL-471 Chapultepec, Méx. Z. mays ssp. mexicana 0.507 3673
JSG-RMM-LCL-477 San Pedro Nexapa, Méx. Z. mays ssp. mexicana 0.464 3746
JSG-RMM-LCL-479 Amecameca, Méx. Z. mays ssp. mexicana 0.491 4105
JSG Y LOS -75 Cojumatlán, Mich. Z. mays ssp. mexicana 0.422 3355
JSG-RMM-LCL-552 Corralitos, Jal. Z. diploperennis 0.423 3651
Agua Blanca Agua Blanca, Guat. Z. luxurians 0.405 3399
Monajil Monajil, Guat. Z. ssp. huehuetenangensis 0.403 3191
114 Martha Isabel Torres-Morán et al.
scientia-CUCBA 9(2):105—117.2007
�����
�����
�����
�����
�����
�����
�����
�����
�����
�����
�����
��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ���
����������������
���������������
5
6
7
8
1
2
3
4
Figura 2. Caracterización espectrofotométrica de ADN de 8 accesiones del género Zea obtenidas por el método Clarke et al. 1989.
�����
�����
�����
�����
�����
�����
�����
�����
�����
�����
�����
��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ���
����������������
���������������
5
6
7
8
1
2
3
4
Figura 3. Caracterización espectrofotométrica de ADN de 8 accesiones del género Zea extraídas por el método de Saghai-Maroof et al. 1984.
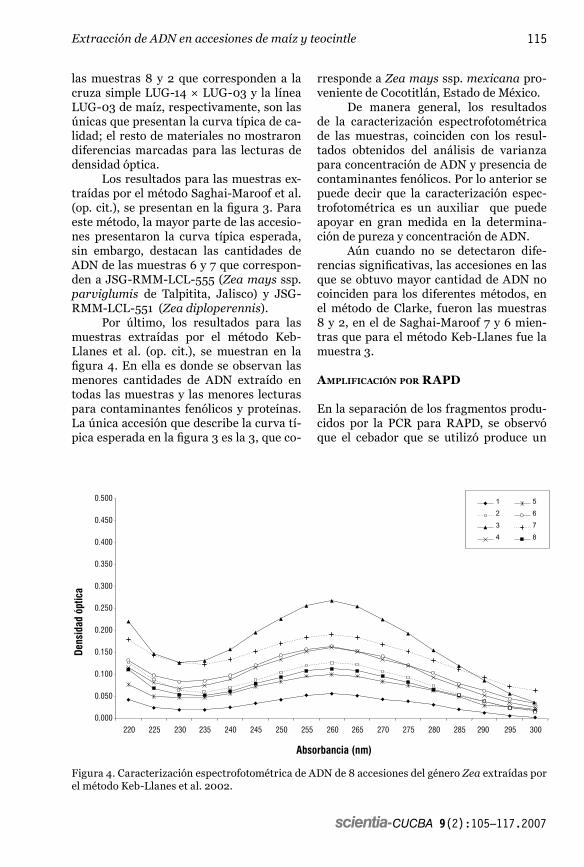
Extracción de ADN en accesiones de maíz y teocintle 115
scientia-CUCBA 9(2):105—117.2007
las muestras 8 y 2 que corresponden a la cruza simple LUG-14 × LUG-03 y la línea LUG-03 de maíz, respectivamente, son las únicas que presentan la curva típica de ca-lidad; el resto de materiales no mostraron diferencias marcadas para las lecturas de densidad óptica.
Los resultados para las muestras ex-traídas por el método Saghai-Maroof et al. (op. cit.), se presentan en la figura 3. Para este método, la mayor parte de las accesio-nes presentaron la curva típica esperada, sin embargo, destacan las cantidades de ADN de las muestras 6 y 7 que correspon-den a JSG-RMM-LCL-555 (Zea mays ssp. parviglumis de Talpitita, Jalisco) y JSG-RMM-LCL-551 (Zea diploperennis).
Por último, los resultados para las muestras extraídas por el método Keb-Llanes et al. (op. cit.), se muestran en la figura 4. En ella es donde se observan las menores cantidades de ADN extraído en todas las muestras y las menores lecturas para contaminantes fenólicos y proteínas. La única accesión que describe la curva tí-pica esperada en la figura 3 es la 3, que co-
rresponde a Zea mays ssp. mexicana pro-veniente de Cocotitlán, Estado de México.
De manera general, los resultados de la caracterización espectrofotométrica de las muestras, coinciden con los resul-tados obtenidos del análisis de varianza para concentración de ADN y presencia de contaminantes fenólicos. Por lo anterior se puede decir que la caracterización espec-trofotométrica es un auxiliar que puede apoyar en gran medida en la determina-ción de pureza y concentración de ADN.
Aún cuando no se detectaron dife-rencias significativas, las accesiones en las que se obtuvo mayor cantidad de ADN no coinciden para los diferentes métodos, en el método de Clarke, fueron las muestras 8 y 2, en el de Saghai-Maroof 7 y 6 mien-tras que para el método Keb-Llanes fue la muestra 3.
AMPLIFICACIÓN POR RAPD
En la separación de los fragmentos produ-cidos por la PCR para RAPD, se observó que el cebador que se utilizó produce un
�����
�����
�����
�����
�����
�����
�����
�����
�����
�����
�����
��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ��� ���
����������������
���������������
5
6
7
8
1
2
3
4
Figura 4. Caracterización espectrofotométrica de ADN de 8 accesiones del género Zea extraídas por el método Keb-Llanes et al. 2002.

116 Martha Isabel Torres-Morán et al.
scientia-CUCBA 9(2):105—117.2007
patrón de amplificación diferente para cada muestra, identificándose algunas bandas polimórficas.
Del gel obtenido, se concluye que se logró una mejor definición de las bandas en las muestras de ADN extractado con el método Keb-LLanes et al. 2002.
Conclusiones
Hubo diferencias marcadas entre los méto-dos de extracción de ADN de Keb-Llanes et al. (op. cit.), Saghai-Maroof et al. (op. cit.) y Clarke et al. (op. cit.), en cuanto a la calidad y concentración de ADN. La mayor concen-tración de ADN se obtuvo con el método de Saghai-Maroof et al. (op. cit.), mientras que la mayor calidad de ADN obtenido fue por el método Keb-Llanes et al. (op. cit.).
Las diferencias entre los métodos se pueden encontrar en varios aspectos de importancia, en particular a la cantidad de reactivos utilizados y el tiempo requerido para obtener el ADN; dichos aspectos son fundamentales para elegir el protocolo. El método Saghai-Maroof et al. (1984) fue el mejor respecto a costos y rapidez.
No se detectaron diferencias en cuan-to a concentración y calidad de ADN entre especies del género Zea ni efectos diferen-ciales debido a los métodos, lo cual indica que los resultados de esta investigación se
pueden usar de manera amplia en el resto de especies y poblaciones del género Zea.
Los diferentes métodos cualitativos y cuantitativos empleados en esta investi-gación, incluyendo la espectrofotometría, fueron consistentes y se complementan como auxiliares en la elección de los pro-tocolos apropiados para la extracción de ADN en el género Zea; podrían requerirse ajustes adicionales dependiendo de los análisis posteriores y de los marcadores moleculares de interés (RAPDs, SSRs, ISTRs, AFLPs).
Literatura citada
Amersham, Biosciences. 2003. Ready-to-go RAPD Analysis Beads Instructions.
http://www.amersham.com Clarke, B.C., L.B. Moran y R. Appels. 1989.
DNA analysis in wheat breeding. Genome 32: 334–339.
Csaikl, U.M., H. Bastian, R. Brettschneider, S. Gauch, A. Meir, M. Schauerte, F. Scholz, C. Sperisen, B. Vornam y B. Ziegenhagen. 1998. Comparative Analysis of Different DNA Extraction Protocols: A Fast, Universal Maxi-Preparation of High Quality Plant DNA for Genetic Evaluation and Phylogenetic Studies. Plant Mol. Biol. Rep. 16: 69–86.
Drábková, L., J. Kirschner y C. Vlcek. 2002. Comparison of Seven DNA Extraction and Amplification Protocols in Historical
MPM300pb A B
Figura 5. Patrones de amplificación de RAPD para 8 accesiones del género Zea. A). Método de Keb-Llanes et al. 2002. B). Método de Saghai-Maroof et al. 1984.

Extracción de ADN en accesiones de maíz y teocintle 117
scientia-CUCBA 9(2):105—117.2007
Herbarium Specimens of Juncaceae. Plant Mol. Biol. Rep. 20: 161–175.
Hillis, D.M., B. K. Mable, A. Larson, S. K. Davis y E. A. Zimmer. 1996. Molecular Systematics. 2a ed. Sinauer Associates, Inc.Sunderland, MA. USA. Pp. 321–344.
Keb-Llanes, M., G. González, B. Chi-Manzanero y D. Infante. 2002. A rapid and simple method for small scale DNA extraction in Agavaceae and other tropical plants. Plant Mol. Biol. Rep. 20: 299.
Lynne, S. P. 1998. The double helix meets the cristal lattice: the power and pitfalls of nu-cleic acid approaches for biomineralogical investigation. American Mineralogist 83: 1593–1607.
Murray, M. G. y J.W. Pitas. 1996. Plant DNA from alcohol-preserved samples. Plant Mol. Biol. Rep. 14: 261–265.
Pandey, R. N., R. P. Adams y L.E. Fluornoy. 1996. Inhibition of Random Amplified Polymorphic DNAs (RAPDs) by plant polysaccharides. Plant Mol Biol Rep. 10: 60–65.
Rogstad, S.H. 2003. Plant DNA Extraction Using Silica. Plant Mol. Biol. Rep. 21: 463– 463.
Saghai-Maroof, M.A., K. M. Soliman, R.A. Jorgensen y R.W. Allard. 1984. Ribosomal DNA spacer-lenght polymor-phisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA. 81: 8014–8018
Valadez, M.E. y G. Kahl. 2000. Huellas de ADN en genomas de plantas (teoría y pro-tocolos de laboratorio). Ed. Mundi-Prensa, México, S.A. de C.V. Pág. 147.
Wolfe, K.A., M.C. Breadmore, J. Ferrance, M.E. Power, J.F. Conroy y P.M. Norris y J.P. Landers. 2002. Electrophoresis 232: 727–733.
FECHA EFECTIVA DE PUBLICACIÓN 29 DE DICIEMBRE DE 2007

118
scientia-CUCBA 9(2):118.2007

scientia-CUCBA 9(2):119—128.2007
Selección por comportamiento per se en líneas de jitomate: producción de fruto y semilla
Eduardo Rodríguez Guzmán1, Aquiles Carballo Carballo2, Gustavo Baca Castillo3, Rogelio Lépiz Ildefonso1, Lino de la Cruz Larios1 y Salvador Mena Munguía1
1Universidad de Guadalajara, CUCBA, IMAREFI. Km. 15.5 Carr. Guadalajara-Nogales, Predio Las Agujas Nextipac, C.P. 44171 Zapopan, Jal. México. Fax 01 33 36-820743.2Colegio de Postgraduados. IREGEP. Km.36.5 Carr. México -Texcoco. C.P. 56230 Montecillo, Estado de México. Tel y Fax. 01(595) 95–20262. 3Colegio de Postgraduados. IRENAT. Km.36.5 Carr. México -Texcoco. C.P. 56230 Montecillo, Estado de México. Tel y Fax.01(595)95 – 20200.Correo electrónico: [email protected].
Resumen: Con el propósito de seleccionar líneas de jitomate por su comportamiento per se para producción de fruto y semilla, y diferenciarlas por su aptitud para participar como progenitores hembra, se estableció un ensayo uniforme en invernadero bajo hidroponía de 25 líneas experimentales, durante primavera-verano de 1995, en Montecillo, México. Se empleo la solución nutritiva universal de Steiner a presión osmótica de 0.7 y 0.9 atm, que resultó con un efecto restringido sobre la producción de fruto y ninguno sobre la producción de semilla. No se presentaron diferencias estadísticas entre líneas en la producción de fruto maduro por planta; en cambio, sí se presentaron diferencias significativas en el número y peso promedio de fruto, así como en los diferentes tamaños en que se clasificó el fruto. Las líneas no difirieron estadísticamente en producción de semilla por planta y en proporción semilla-fruto; sin embargo, en número y peso promedio de semillas por fruto se obtuvieron diferencias significativas. Se encontraron correlaciones positivas entre producción con número y peso promedio de fruto por planta, y entre producción de semilla por planta con producción de fruto y número de frutos por planta. El comportamiento per se de las líneas para los componentes de producción de fruto y semilla hizo posible seleccionar las sobresalientes con base a número, peso promedio y tamaño de fruto; y número y peso promedio de semilla por fruto. Las líneas identificadas como sobresalientes integrando características de fruto y semilla fueron: CPJL-21-1 y CPJL-24-5 en el grupo 1; CPJL-8-7 en el grupo 2; y en el grupo 3 CPJL-1, CPJL-22-2.
Palabras clave: Lycopersicon esculentum Mill.; mejoramiento genético; selección; producción de semilla.
Abstract: A uniform trial was carried out during summer 1995 under greenhouse and hydroponics conditions at Montecillo, México. The objective was to select tomato lines by their per se performance for fruit and seed production, and then differentiate them by their aptitude for female parent. Was compared 25 tomato experimental lines under two nutrient solutions with different osmotic pressure. Was not differences between solutions for seed production and only extra big size of fruit tomato was the only one character affect of fruit. Tomato lines were similar in mature fruit production per plant, but number and fruit weight per plant, and all the classifications of tomato size presented significant differences. The lines were similar in seed production per plant and fruit–seed proportion, but differences were obtained for seed number and weight average per plant. Positive correlations were found between fruit production with number and weight average of fruit per plant; and between seed production per plant with fruit production and number of fruits per plant. Per se performance of tomato lines for fruit and seed production components made possible to select the top lines, the most useful characters

120 Eduardo Rodríguez Guzmán et al.
scientia-CUCBA 9(2):119—128.2007
to the selection were number, weight average and size for fruit, and number and weight average of seed per fruit. The selected top lines with good fruit and seed production were CPJL-21-1 y CPJL-24-5 en el grupo 1; CPJL-8-7 en el grupo 2; y en el grupo 3 CPJL-1, CPJL-22-2.Key words: Lycopersicon esculentum Mill.; tomato breeding; selection, seed produc-tion.
Introducción
El mejoramiento genético tiene como pro-pósito incorporar en una variedad las ca-racterísticas favorables de sus progenitores (Márquez-Celis 1988). Varios investigado-res han explorado técnicas por las cuales la superioridad de progenies pudiera prede-cirse, lo que habilitaría al fitomejorador a concentrar esfuerzos sólo en los materiales prometedores desde los primeros ciclos del mejoramiento.
Martínez (1988) menciona que en maíz las variedades con los rendimientos más altos producen las mejores progenies F1 por lo que sugirió para identificar líneas progenitoras superiores, la selección por rendimiento per se. Por su parte, Falconer (1978) afirmó que si el comportamiento de las líneas es función de su contenido gené-tico aditivo, el mejor método para identi-ficar las sobresalientes es probarlas como tales, de esta manera se evita el enmasca-ramiento de genes, se optimiza la varianza genética aditiva entre líneas y se anula la interacción línea por probador.
Con el advenimiento de los híbri-dos de cruza simple, los mejoradores de maíz han puesto mayor interés en el me-joramiento del rendimiento per se (por sí mismo) de las líneas, al considerar que el mejor método para formar líneas y cruzas simples de alto rendimiento, es la evalua-ción de ambas como tales (Martínez-Celis op. cit.).
Este método para seleccionar líneas altamente rendidoras fue propuesto por Conti et al. (1990) para discriminar entre líneas progenitoras de jitomate para ren-dimiento comercial y total de fruto, y pro-ducción de sólidos solubles. Peirce (1991) señaló que la aptitud combinatoria gene-
ral (ACG) y los valores promedio de los progenitores correlacionaron bien con el promedio de familias derivadas por selec-ción uniseminal en jitomate; en tanto que Dudi y colaboradores (citados por Kalloo 1988) observaron una estrecha asociación entre el comportamiento medio de los pro-genitores y el de los híbridos con aptitud combinatoria general y específica para ren-dimiento y sus componentes.
Oliveira (citado por Nascimento et al. 1994) señaló que en las especies hor-tícolas las variedades con una reducida producción de semilla representan para las empresas de semillas un mayor costo de producción, por lo que esta caracterís-tica asume una importancia relativa como criterio de selección en los programas de mejoramiento y vuelve prioritario identifi-car y seleccionar materiales genéticos que simultáneamente presenten las caracte-rísticas de interés para la agroindustria y para las empresas productoras de semillas.
Hernández et al. (1992), al estudiar las características para producción de se-milla básica de seis variedades de jitomate, encontraron que las variedades con alto rendimiento en frutos, obtuvieron bajo rendimiento en semilla o viceversa, con rendimientos de semilla por planta que va-riaron de 1.88 a 3.48 gramos por planta y señalaron que estos aspectos son influidos por características varietales heredables y por las condiciones ambientales.
En el presente estudio se planteó inferir la selección de líneas de jitomate como progenitores hembra en base a su comportamiento per se en producción de fruto y semilla, y valorar la influencia del

Selección por comportamiento per se en líneas de jitomate... 121
scientia-CUCBA 9(2):119—128.2007
ambiente sobre la expresión de este com-portamiento.
Materiales y métodos
El estudio se realizó en las instalaciones del Programa Interdisciplinario de Producción de Semillas, adscrito al Instituto de Recursos Genéticos y Productividad del Colegio de Posgraduados, Montecillo, Estado de México, durante primavera-verano de 1995. El trabajo experimental consistió de un ensayo bajo hidroponía para estimar producción de fruto y semilla de 25 líneas experimentales de jitomate, que se obtuvieron del Proyecto de Jitomate en el área de Mejoramiento y Control de la Calidad Genética (cuadro 1). El ensayo se estableció en invernadero, se utilizó la so-lución nutritiva universal de Steiner a pre-siones osmóticas de 0.7 y 0.9 atm, con pH en 5.5, y las relaciones mutuas universales de aniones y cationes: K+ : Ca++ : Mg++ = 35: 45 : 20 y NO3- : H2PO4- : SO4-- = 60 : 5 : 35 (Steiner 1984). En grado comercial se emplearon los fertilizantes: sulfato de mag-nesio, nitrato de calcio y nitrato de potasio; y como reactivos analíticos Monterrey®: fosfato de potasio y sulfato de potasio. Dado el alto grado de pureza observado en los fertilizantes, para los cálculos y prepa-ración de las soluciones se empleo el peso molecular de reactivos grado analítico.
Se descontaron los nutrimentos en-contrados en muestreos de agua de riego que se realizaron el 2 de junio, para pre-parar solución nutritiva en uso desde el trasplante, y el 20 de septiembre de 1995,
para la solución en uso desde esta fecha hasta finalizar la cosecha, la cual se realizó cuando el fruto se observó maduro por su consistencia y color. Se utilizó un sistema hidropónico abierto consistente en 4 camas de siembra de 9.80 m de largo, 0.50 m de ancho y 0.20 m de profundidad, distancia-das entre sí 0.75 m; el substrato fue tezon-tle rojo granulometría 2-10 mm. Se sembró el 29 de mayo en charolas de poliestireno con arenilla de tezontle, y se trasplantó el 2 de agosto; la cosecha abarcó de noviembre 7 de 1995 a enero 6 de 1996.
El diseño experimental fue bloques completos al azar y arreglo en parcelas divididas con dos repeticiones, cada una estuvo representada por una cama de siembra con dos hileras de plantas sepa-radas entre sí 30 cm y 35 cm entre plan-tas, para tener un total de 50 plantas por cama. Como parcela grande se probaron soluciones nutritivas y como subparcela genotipos, la unidad experimental se con-formo por 2 plantas. Las variables para fruto maduro fueron: producción por planta, producción por tamaños (cuadro 2) y peso promedio en gramos, y número de frutos por planta. Para semilla se conside-ró producción de semilla por planta y peso promedio de semillas por fruto en gramos, número de semillas por fruto, y proporción semilla-fruto. Con los resultados se realizó análisis de varianza y comparación de me-dias mediante la prueba de Tukey al 5% de probabilidad, se estimaron correlaciones entre variables de producción de fruto y semilla.
Cuadro 1. Relación de genotipos de jitomate empleados.
Líneas experimentales
1 CPJL-1 6 CPJL-7-8 11 CPJL-18-8 16 CPJL-22-4 21 CPJL-25-14
2 CPJL-2 7 CPJL-8-7 12 CPJL-19-1 17 CPJL_22-7 22 CPJL-26-8
3 CPJL-3 8 CPJL-9-2 13 CPJL-20-9 18 CPJL-23-4 23 CPJL-27-9
4 CPJL-4-8 9 CPJL-10-1 14 CPJL-21-1 19 CPJL-24-5 24 CPJL-28-7
5 CPJL-5-4 10 CPJL-17-7 15 CPJL-22-2 20 CPJL-25-8 25 CPJL-28-11

122 Eduardo Rodríguez Guzmán et al.
scientia-CUCBA 9(2):119—128.2007
Resultados y discusión
Entre soluciones nutritivas no se obtuvieron diferencias significativas en ninguno de los caracteres estudiados, mientras que entre genotipos se manifes-taron diferencias significativas en número de frutos por planta y peso promedio por fruto. La interacción entre soluciones y genotipos en ninguna variable resulto sig-nificativa (cuadro 3).
En producción de fruto maduro por tamaños se manifestaron diferencias signi-ficativas (P ≤ 0.05) entre soluciones nutri-tivas únicamente para el fruto extra gran-de; entre genotipos en todos los tamaños se obtuvieron diferencias. Tampoco existió interacción significativa entre soluciones y genotipos en ningún tamaño (cuadro 4). En relación a las soluciones nutritivas, los me-jores resultados para tamaño extragrande de fruto maduro se obtuvieron bajo la so-lución a 0.7 atm, con promedio de 571.97 g, contra 402.12 g bajo la solución con 0.9 atm. Entre los genotipos, por número de
Cuadro 2. Clasificación por tamaño de fruto estimado de acuerdo al diámetro ecuatorial. Normas americanas de calidad para tomate fresco, León y Arosemena (1980).
Tamaño Mínimo (mm)
Máximo (mm)
1. Rezaga* <35
2. Rezaga* 35 48
3. Extrachico (7x8) 48 54
4. Chico (7x7) 54 58
5. Mediano (6x7) 58 64
6. Grande (6x6) 64 73
7. Extragrande (5x5) (5x6) 73 88
8. Máximo grande (4x5) >88
*Se anexo un tamaño en rezaga para diferenciar frutos en los jitomates tipo saladette, por lo que la categoría rezaga 2 no es fruto de desecho y exclusivamente la rezaga tipo 1 sería la que no cumplió las características deseables para su comercialización.
frutos destacaron las líneas 14, 3, 20 y 21 de fruto tipo saladet, excepto la línea 3 que produce fruto tipo bola. Por peso promedio de fruto sobresalieron los genotipos 1, 15 y 16 con fruto tipo bola (cuadro 5).
La correlación entre número y peso promedio de fruto maduro, con producción de fruto maduro por planta fue de 0.55 y 0. 26, respectivamente, lo que indica que la asociación entre producción de fruto maduro por planta con el número de fru-tos por planta es más estrecha que con el peso promedio por fruto, en tanto que la correlación entre número de frutos y peso promedio fue significativa, pero en sentido negativo (-0.58). Al separar la producción de fruto maduro por planta por tamaños, es posible agrupar los diferentes genotipos en tres clases: Grupo 1: líneas con produc-ción de frutos de tamaños denominado rezaga 2 a mediano (figura 1a), Grupo 2: líneas con producción de frutos de tama-ños extrachico a grande (figura 1b), Grupo 3: líneas con producción de frutos de ta-maños mediano a extragrande (figura 1c). Con base en esta agrupación y en el rendi-miento total de fruto es posible seleccionar dentro del grupo 1 la línea 14 como una de las más productivas, principalmente en los tamaños rezaga 2 y extrachico de fruto tipo saladet (figura 1a); dentro del grupo 2 con fruto tipo bola sobresalen los genotipos 7 y 18 con los valores más altos de producción por planta (cuadro 5), la primera línea con mayor producción en fruto extrachico a mediano y la segunda con producción de fruto de tamaños mediano a grande (figura 1b); y dentro del grupo 3 de fruto tipo bola con mejor producción de fruto de tamaños grande y extragrande, los genotipos 1 y 15 (figura 1c).
Respecto a la producción de semilla, no se presentaron diferencias significa-tivas entre las soluciones nutritivas para las variables en estudio. Entre genotipos, en producción de semilla por planta y en proporción semilla-fruto no se registraron diferencias significativas, mientras que en número y peso promedio de semilla por fruto si se presentaron diferencias esta-

Selección por comportamiento per se en líneas de jitomate... 123
scientia-CUCBA 9(2):119—128.2007
Cuadro 3. Cuadrados medios de los análisis de varianza para 6 caracteres de producción de fruto en jitomate.
Fruto maduro Semilla
FV GL Producción por planta
Número por
planta
Peso promedio por fruto
Producción por planta
Proporciónsemilla
fruto
Número por fruto
Peso promedio por fruto
Rep 1 862000 259 69 0.43 0.62 195010 0.026
Sol 1 324580 72 1612 0.03 0.31 2420 0.002
Ea 1 172756 127 102 2.13 0.48 333853 0.019
Gen 24 320871 183 ** 1624 ** 3.51 0.23 200855 ** 0.012 **
SxG 24 349284 49 113 5.37 0.14 61836 0.005
Eb 48 362276 55 140 5.35 0.16 58911 0.004
CV 24 23 4 24.28 12.18 24.55 24.77
*, ** Significativo a una P≤0.05, 0.01, respectivamente. FV=Fuente de variación; GL=Grados de Libertad; Rep=Repetición; Sol=Solución; Ea=Error a; Gen=Genotipos; SxG=Interacción entre soluciones y genotipos; Error b; CV=Coeficiente de variación.
Cuadro 4. Cuadrados medios de los análisis de varianza para la producción de fruto maduro por tamaños (1 a 8) en jitomate.
FV GL Producción por planta de fruto maduro por tamaños
Rezaga
1
Rezaga 2 Extra chico Chico Mediano Grande Extra
grande
Máximo
grande
Rep 1 8502 92148 41734 98 289508 5915 19132 12588
Sol 1 1369 169184 426892 16 94802 1344973 98420 * 9801
Ea 1 3892 58264 5887 3014 12800 24532 170 10882
Gen 24 3513 ** 597258 ** 306823 ** 196944 ** 287234 ** 347266 ** 148225 ** 21112 **
SxG 24 898 33757 60865 47865 93216 88640 32940 4872
Eb 48 963 48976 82831 59017 99446 99665 24467 5901
*, ** Significativo a una P ≤ 0.05, 0.01, respectivamente. FV=Fuente de variación; GL=Grados de Libertad; Rep=Repetición; Sol=Solución; Ea=Error a; Gen=Genotipos; SxG=Interacción entre soluciones y genotipos; Eb=Error b.
dísticamente significativas (P ≤ 0.01). En la interacción entre genotipos y soluciones nutritivas no se manifestó significancia es-tadística (cuadro 3).
La línea 15 estadísticamente superior al resto en número y peso promedio de semillas por fruto, fue seguida por la línea 1; por el contrario las líneas 3, 14, 20 y 21 produjeron las menores cantidades y pesos
promedio de semilla por fruto, principal-mente en tamaños rezaga 2 y extra chico (cuadro 5).
La producción de semilla por planta correlacionó de manera significativa y po-sitiva con el rendimiento de fruto de los tamaños extrachico (0.352), chico (0.397) y mediano (0.251), mientras que con la producción de fruto de tamaño máximo

124 Eduardo Rodríguez Guzmán et al.
scientia-CUCBA 9(2):119—128.2007
Cuadro 5. Medias de 25 genotipos de jitomate para variables de producción de fruto maduro. Montecillo, México. 1995-1996.
Geno-
tipo
Fruto maduro Semilla
Produc-
ción por
planta
Frutos por
planta
Peso promedio por
fruto
Produc-
ción por
planta
Propor-
ción
semilla-
fruto
Número por
fruto
Peso
promedio
por fruto
1 2543.4 18.75 g 138.83 a 6.60 2.91 1334 B 0.36 b
2 2155.5 25.75 fg 83.80 bcdefg 7.95 3.49 1089 bcdef 0.31 bcd
3 2586.4 44.75 ab 60.68 jkl 7.68 3.13 647 H 0.17 f
4 2463.6 35.00 bcdef 70.10 fghijk 8.53 3.33 858 cdefgh 0.24 cdef
5 2379.1 31.00 def 76.38 efghijk 8.50 3.47 984 cdefgh 0.28 bcde
6 2679.7 30.00 ef 89.98 bcde 9.08 3.32 1098 bcdef 0.30 bcd
7 3052.4 33.50 cdef 90.75 bcde 9.05 3.13 982 cdefgh 0.27 bcde
8 2629.2 33.75 cdef 78.48 cdefghi 10.53 3.64 1094 bcdef 0.31 bcd
9 2575.6 30.00 ef 83.80 bcdef 7.90 3.15 889 cdefgh 0.26 cdef
10 2639.4 32.75 cdef 80.18 cdefgh 8.33 3.26 823 defgh 0.26 cdef
11 2518.5 37.25 abcde 67.80 ghijk 9.80 3.56 942 cdefgh 0.27 bcde
12 2452.6 26.25 fg 93.60 cbd 8.60 3.40 1130 bcde 0.33 bc
13 2409.4 36.00 abcdef 66.65 hijkl 8.55 3.39 851 cdefgh 0.23 def
14 2781.1 45.75 a 60.40 kl 8.63 3.20 716 gh 0.19 ef
15 2361.9 18.50 g 129.93 a 8.28 3.35 1731 A 0.46 a
16 2932.3 29.50 ef 99.98 b 6.53 2.66 784 fgh 0.23 def
17 2389.1 30.75 def 77.45 defghij 7.83 3.31 928 cdefgh 0.26 cdef
18 2694.6 28.75 efg 94.35 cb 8.80 3.29 1148 bcd 0.31 bcd
19 2435.1 32.00 cdef 75.75 efghijk 7.73 3.22 881 cdefgh 0.24 cdef
20 2493.7 41.00 abcd 61.93 ijkl 8.95 3.42 799 efgh 0.22 def
21 2056.9 41.75 abc 49.85 l 8.08 3.59 803 defgh 0.25 cdef
22 2601.0 32.00 cdef 79.38 cdefgh 9.28 3.48 1017 bcdefg 0.29 bcd
23 2407.5 27.75 efg 86.33 bcdef 6.83 3.09 989 bcdefgh 0.25 cdef
24 2030.0 26.25 fg 74.15 efghijk 7.43 3.53 1186 bc 0.29 bcd
25 1708.3 29.50 ef 63.80 hijkl 7.60 3.81 1101 bcdef 0.28 bcde
Medias con letras iguales no son estadísticamente diferentes (Tukey, 0.05)
grande fue significativa pero con sentido negativo (-0.224).
Entre las variables producción de semilla por planta con producción de fruto maduro por planta y número de frutos por
planta, la correlación fue significativa y positiva (0.52 y 0.44, respectivamente), en tanto que con peso promedio de fruto por planta no fue significativa y fue de signo negativo (-0.05).
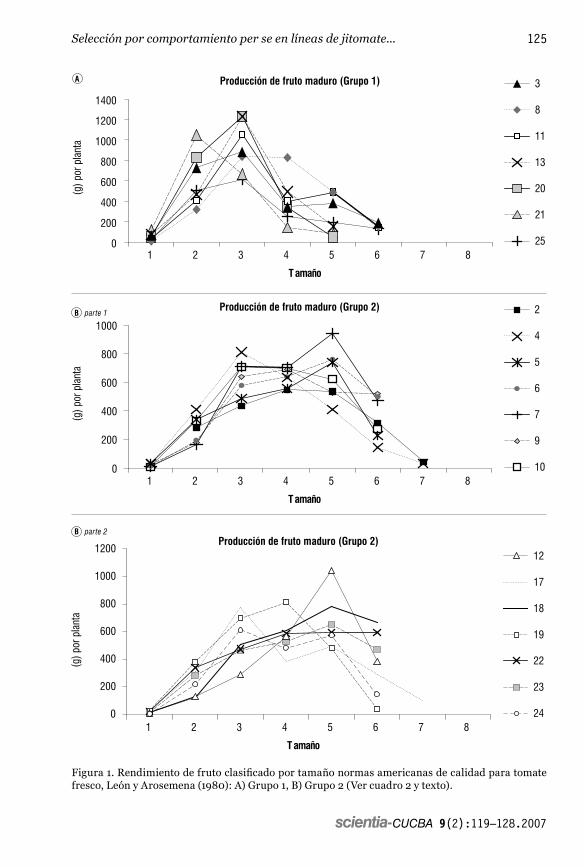
Selección por comportamiento per se en líneas de jitomate... 125
scientia-CUCBA 9(2):119—128.2007
�������������������������������������
� � � � � � � �
������
��������������
�
�
��
��
��
��
���
���
���
���
���
����
����
����
A
�������������������������������������
� � � � � � � �
������
��������������
�
�
�
�
�
�
���
���
���
���
���
����B parte 1
�������������������������������������
� � � � � � � �
������
��������������
��
��
��
��
��
��
���
����
����
���
���
���
���
Figura 1. Rendimiento de fruto clasificado por tamaño normas americanas de calidad para tomate fresco, León y Arosemena (1980): A) Grupo 1, B) Grupo 2 (Ver cuadro 2 y texto).
B parte 2

126 Eduardo Rodríguez Guzmán et al.
scientia-CUCBA 9(2):119—128.2007
Las presiones osmóticas de las so-luciones nutritivas utilizadas tuvieron un efecto limitado sobre la producción de fru-to y ninguno sobre la producción de semi-lla, lo que no concuerda con lo planteado por Steiner (citado por Baca 1985) quién consideró a la presión osmótica de la solu-ción nutritiva como el mayor determinante del crecimiento, desarrollo y producción de la planta, y su efecto dependiente de la clase de planta y del clima, dado que era de esperarse que se presentaran diferencias entre soluciones que además interacciona-ran con los genotipos.
La respuesta a las soluciones nutri-tivas de la producción de fruto de tamaño extragrande podría estar relacionada con el hábito de crecimiento, pues las plantas que produjeron frutos de este tipo fueron de hábito indeterminado y fruto tipo bola. Rodríguez (1996) al comparar el compor-tamiento entre una variedad de hábito determinado y otra de hábito indetermi-nado encontró que en la primera no hubo deferencias en la respuesta a diferentes niveles de presión osmótica, mientras que la última mostró mejor comportamiento bajo una presión osmótica de 0.7 atm, con reducción en su producción a mayor o me-nor presión osmótica.
Las producciones de semilla por planta bajo hidroponía son mayores a al-gunas producciones obtenidas en campo, como en el presente estudio donde los pro-medios variaron entre 6.60 g y 10.53 gra-mos; Martínez-Loeza (1992) obtuvo 14.8 gramos en una variedad y 19.4 gramos en un híbrido, en tanto que Hernández et al. (1992) reportan bajo condiciones de campo rendimientos de semilla por planta que van desde 1.88 a 3.48 gramos por planta. Es de interés estimar si existen ventajas en la producción de semilla bajo invernadero e hidroponía, aún para semillas que serán utilizadas en campo por una posible ven-taja en la calidad de la semilla, además de una mayor producción.
Los genotipos que se estudiaron tuvieron similar capacidad productiva de fruto maduro por planta, resultando de interés la presencia de diferencias signifi-cativas en el número y peso promedio de fruto maduro, y en los diferentes tamaños en que se clasifico el fruto. La similitud en producción de fruto maduro entre los 25 genotipos se pudo deber a que las plantas compensaron el mayor número de frutos de menor tamaño, y viceversa. Este com-portamiento se reforzó con los resultados del análisis de correlación, donde número
�������������������������������������
� � � � � � � �
������
��������������
�
��
��
�
����
����
���
���
���
���
Figura 1. Continuación. Rendimiento de fruto clasificado por tamaño normas americanas de cali-dad para tomate fresco, León y Arosemena (1980): C) Grupo 3 (Ver cuadro 2 y texto).
C

Selección por comportamiento per se en líneas de jitomate... 127
scientia-CUCBA 9(2):119—128.2007
y peso de fruto mostraron valores positivos con producción de fruto maduro por plan-ta, más estrecha la relación con el número y menos con el peso promedio de fruto. La correlación entre número y peso promedio de frutos significativa pero con sentido ne-gativo (-0.58) indica que entre ambos ca-racteres existe un efecto de competencia.
Narendra y Arya (1995) en 34 geno-tipos de jitomate encontraron que número de frutos por planta mostró asociación positiva con rendimiento de frutos por planta, carácter que más contribuyó al ren-dimiento. Moya et al. (1996), encontraron que el número de frutos por planta fue el carácter con mayor efecto directo sobre rendimiento en tres grupos de variedades: de crecimiento indeterminado de fruto grande, de crecimiento determinado de fruto grande y de crecimiento determinado de fruto chico. El peso de fruto correlacio-nó significativamente con signo positivo, con el rendimiento en las variedades de-terminadas de fruto grande; y con signo negativo en las variedades de fruto chico.
La asociación negativa entre número y peso de fruto puede ser propiciada por limitaciones genéticas de la capacidad de producir fruto, como lo señala Griffing (citado por Singh et al. 1974). Este autor propuso que estos caracteres están deter-minados por el mismo complejo de genes cuya principal función parece ser controlar su balance.
En la producción de semilla por plan-ta y en la proporción semilla-fruto no se presentaron diferencias significativas en-tre los 25 genotipos empleados. Martínez-Loeza (op. cit.) en condiciones similares comparó la producción de semilla entre la variedad de polinización libre Floradade y el híbrido (Contessa), y tampoco registró diferencias entre variedades para estas características.
Las diferencias obtenidas entre las 25 líneas en número y peso promedio de semi-lla por fruto, pudieron presentar un efecto compensatorio con el número o tamaño de los frutos, por lo que en líneas con mayor cantidad de fruto por planta, estos fueran
de menor tamaño y con menor cantidad de semilla; mientras que líneas con me-nor cantidad de frutos de mayor tamaño y peso, tuvieran mayor cantidad de semilla por fruto; esto podría explicar la similitud entre líneas en producción de semilla por planta. La correlación en producción de semilla por planta con producción de fruto maduro por planta y número de frutos por planta fue alta y positiva, y más asociada a la producción de frutos de tamaño chico a mediano, a causa del número mayor de frutos por planta.
Las variedades con una reducida producción de semilla representan para las empresas de semillas mayor costo de pro-ducción (Oliveira, citado por Nascimento et al. op. cit.), y resulta prioritario iden-tificar y seleccionar materiales genéticos que simultáneamente presenten las carac-terísticas de interés para la agroindustria y las empresas productoras de semillas (Nascimento et al. op. cit.); por lo tanto, las líneas progenitores hembra deben ser plantas que tengan una mejor producción de semilla por planta. Por esta razón se recomienda al fitomejorador que incluya como criterios en un programa de mejora-miento genético el número y peso prome-dio de fruto por planta, así como el número y peso de semilla por fruto, de tal manera que le permitan seleccionar como progeni-tores hembra aquellos genotipos que pre-senten valores mayores para los caracteres mencionados, que a la vez compensen los efectos negativos producidos entre sí.
Conclusiones
La selección de líneas de jitomate como progenitores hembra en base a su com-portamiento per se en producción de fruto y semilla esta en función de una relación presente entre el tamaño o peso y el núme-ro de frutos, que se asocia a la producción y número de semillas por fruto, donde a mayor tamaño de fruto mayor cantidad de semilla y viceversa, pero también a mayor cantidad de semilla, se obtiene semilla de menor peso.

128 Eduardo Rodríguez Guzmán et al.
scientia-CUCBA 9(2):119—128.2007
Sin embargo, existe la posibilidad de seleccionar genotipos que presentan buen rendimiento de fruto de tamaño aceptable comercialmente y que a su vez producen una cantidad aceptable de semilla como la variedad CPJL-8-7.
La influencia del ambiente, represen-tado por los niveles de presión osmótica de la solución nutritiva, sobre la expresión de las características de producción de fruto y semilla sólo manifestó una ligera influen-cia en el rendimiento de fruto de tamaño extragrande.
Las principales características de fruto que permitieron la selección de lí-neas fueron el número, peso promedio y tamaño; mientras que en semilla fueron el número y peso promedio por fruto.
Bibliografía
Baca-Castillo, G. A. 1983. Efecto de la solu-ción nutritiva, la frecuencia de riegos, el substrato y la densidad de siembra en culti-vos hidropónicos al aire libre de pepino, me-lón y jitomate. Tesis de Doctor en Ciencias. Colegio de Postgraduados, Chapingo, México. 240 pp.
Conti, S., M. C. Sanguineti and R. Roncarati. 1990. Hybrid performance as compared to parents in processing tomato. Adv. in Hort. Sci. 4: 151–154.
Falconer, D. S. 1978. Introducción a la genéti-ca cuantitativa. Traducción del Inglés por F. Márquez S. Ed. CECSA. México. 430 pp.
Hernández, A., R. Díaz, T. Sánchez y M. Rosales. 1992. Características de seis va-riedades de tomate para la producción de semilla básica. Agrotecnia de Cuba 24: 39–45.
Kalloo, G. 1988. Vegetable breeding (V I). CRC Press. Florida, USA. pp. 41–43.
León-Gallegos, H. M. y M. Arosemena-Butari. 1980. El cultivo del tomate para consumo fresco en el valle de Culiacán. SARH-INIA. México. 80 pp.
Márquez-Sánches, F. 1988. Genotecnia Vegetal. Tomo II. AGT. México. 665 pp.
Martínez-Celis, J. J. 1988. Mejoramiento convergente en líneas de maíz conside-rando rendimiento, adaptabilidad y ca-lidad de semilla como criterios de selec-ción. Tesis Maestro en Ciencias. Colegio de Postgraduados, Montecillo, México. 140 pp.
Martínez-Loeza, A. 1992. Producción de ji-tomate bajo hidroponia en condiciones de invernadero. Tesis de Maestro en Ciencias. Colegio de Postgraduados, Montecillo, México. 113 pp.
Moya, C, M. Álvarez, M. Varela y M. E. Mesa. 1996. Estimaciones de parámetros genético-estadísticos en tres grupos diferen-tes de variedades de tomate (Lycopersicon esculentum Mill.) cultivadas en organopó-nico. Cultivos Tropicales 17: 67–71.
Narendra, K. and M.C. Arya. 1995. Association among yield components in tomato. Madras Agricultural Journal 82: 536–539.
Nascimento, W. M., J. B. C. Silva, H. B. S. V. Pessoa. 1994 Produçao de semen-tes de tomate para industria em Brasilia. Horticultura brasileira 12: 96–198.
Peirce, L. C. 1977. Impact of single seed des-cent in selecting for fruit size, earliness and total yield in tomato. Journal of American Society of Horticultural Science 102: 520–522
Rodríguez-Guzmán, E. 1996. Evaluación y selección de líneas como progenitores para la producción de híbridos de jitomate. Tesis de Maestría en Ciencias. Especialidad en Producción de semillas. Colegio de Postgraduados en Ciencias Agrícolas, Montecillo, México. 124 pp.
Singh, H. N., R. R. Singh, and R. K. Mital. 1974. Genotypic and phenotypic variability in tomato. Indian Journal of Agricultural Science 44: 807–811.
Steiner, A. A. 1984. The Universal Nutrient Solution. Sixth Int. Cong. on Soiless Culture ISOSC Preoceeding. The Netherlands. pp: 633–649.
FECHA EFECTIVA DE PUBLICACIÓN 29 DE DICIEMBRE DE 2007

scientia-CUCBA 9(2):129—141.2007
El factor gametofítico-1 (ga1) en teocintle (Zea spp.) en México
Lino De la Cruz Larios, José de Jesús Sánchez González, José Guadalupe Rodríguez Flores, José Ron Parra y Fernando Santacruz-RuvalcabaUniversidad de Guadalajara, Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA), Departamento de Producción Agrícola, Instituto de Manejo y Aprovechamiento de Recursos Fitogenéticos (IMAREFI), Km 15.5 Carretera Guadalajara-Nogales. CP 45110. Apartado Postal 129, Las Agujas, Nextipac, Mpio de Zapopan, Jalisco, México Tel. y Fax: 33 36 82 07 43.
Resumen: Este estudio se llevó a cabo con la finalidad de conocer la composición alélica del locus ga1 de las especies silvestres de Zea. Los materiales genéticos utilizados para llevar a cabo pruebas de incompatibilidad fueron 60 accesiones de teocintle representativas de la diversidad conocida en México. Los probadores usados involucraron líneas de la Universidad Estatal de Carolina del Norte y de la Universidad de Wisconsin, homocigóticas para el alelo Ga1-S; además se usó un híbrido comercial recesivo para ga1. Se caracterizaron 60 accesiones de teocintle y las diferencias en la formación de grano entre accesiones y grupos de accesiones fueron determinadas con base en los procedimientos GLM y MIXED del Sistema de Análisis Estadístico (SAS). El alelo Ga1-S, responsable de la incompatibilidad no recíproca fue detectado en el 57% de las accesiones de teocintle; por su parte el alelo Ga1-m, compatible con cualquier otro genotipo del locus ga1, se presentó únicamente en 25% de las accesiones mientras que un 18% de las poblaciones fueron homocigóticas ga1/ga1.
Abstract: This study was carried out in order to know the allelic composition of the locus ga1 in the wild species of Zea. Genetic materials used for the incompatibility tests were 60 teosinte accessions representing the diversity found in Mexico. The testers used introduced inbred lines from North Carolina State University and the University of Wisconsin: homozygous for Ga1-S allele. In addition, a hybrid recessive for ga1 was used. Sixty teosinte accessions were characterized, and the differences in the seed set among accessions and groups of accessions were determined based on GLM and MIXED procedures of the Statistical Analysis System (SAS). The allele Ga1-S, responsible for non-reciprocal incompatibility, was detected in 57% of the teosinte accessions. Ga1-m allele, on the other hand, which is compatible with any other allele of locus ga1, was present only in 25% of the accessions, while 18% of the populations were homozygous ga1/ga1.
Introducción
Durante la última década ha existido un interés creciente para entender los diferen-tes factores involucrados en el flujo genéti-co entre especies y poblaciones de plantas. Entre los aspectos de mayor importancia que motivan el interés en los estudios del flujo genético en México, se pueden men-cionar a la posible incorporación del maíz transgénico a la producción agrícola na-cional y su impacto en las variedades na-tivas y especies silvestres emparentadas,
el mantenimiento de la pureza varietal en la producción de semilla para siembra y la producción comercial de variedades de maíz de usos especiales libres de contami-nación por variedades diferentes.
Después de casi un siglo de estudios detallados, se acepta ampliamente que la domesticación del maíz (Zea mays L.) ocu-rrió en México hace alrededor de 10,000 años a partir de una especie tropical de teocintle anual, Zea mays ssp. parviglu-

130 Lino De la Cruz Larios et al.
scientia-CUCBA 9(2):129—141.2007
mis Iltis & Doebley (Matsuoka et al. 2002; Doebley 2004). Los parientes silvestres del maíz, de manera colectiva llamados “teo-cintle”, están representados por especies anuales, perennes diploides y tetraploides. La presencia de híbridos entre el maíz y el teocintle está bien documentada y hay evi-dencias directas en los registros arqueo-lógicos de que han estado intercambiando genes por mucho tiempo (Mangelsdorf et al. 1967 y Wilkes 1977). Sin embargo, va-rios investigadores señalan que el flujo ge-nético entre maíz y teocintle es un evento más raro de lo que podría esperarse, y que esto es debido a varios factores, entre los que destacan los mecanismos de incompa-tibilidad.
Las bases científicas de la incom-patibilidad entre plantas de polinización cruzada como las especies del género Zea, están en un nivel bajo de entendimiento. Nelson (1996) y Kermicle (2006) estudia-ron los factores de incompatibilidad más importantes en maíz, los que se refirieron colectivamente como factores gametofíti-cos. Éstos son numerosos y fueron ubica-dos para los cromosomas 1 (ga4 y ga6), 3 (ga7), 4 (ga1, tcb1), 5 (ga2 y ga10), 7 (ga3), y 9 (ga8). El locus gametofítico identifi-cado como ga1, ha sido sujeto de diversos estudios por cerca de un siglo. Este locus, situado en el brazo corto del cromosoma 4, es el más estudiado; se considera impor-tante a causa de que el alelo Ga1-S condi-ciona esterilidad no-recíproca con el alelo ga1.
Por su parte los factores que limi-tan el cruzamiento del maíz con el teocin-tle se han designado de manera amplia como “Complejo de Incompatibilidad del Teocintle” (TIC). Cuando se lleva polen de teocintle a estigmas de maíz, se obtienen progenies híbridas sin ninguna dificultad; sin embargo, las cruzas recíprocas, es decir cuando el polen de maíz se deposita en los estigmas de teocintle, no siempre tiene éxi-to en la polinización. Por ejemplo, plantas del teocintle Zea mays ssp. mexicana (teo-cintle tipo maleza de El Valle de México y de El Bajío) polinizadas con polen de maíz
presentan una formación de grano irregu-lar, por que no se logra la fecundación en todos los casos. También, cuando se utiliza polen de maíz, en estigmas de teocintle Zea mays ssp. parviglumis (teocintle de las partes bajas de México que crece pre-dominantemente silvestre), la formación de grano es por lo general buena. Evans y Kermicle (2001) describieron el locus Tcb1 (Teosinte crossing barrier-1), responsable del reconocimiento entre el polen y los estigmas, el cual “podría estar desem-peñando un papel muy importante en el aislamiento reproductivo entre el maíz y el teocintle”.
Aún cuando se dispone de literatura relacionada con los sistemas de incom-patibilidad en maíz y teocintle, la mayor parte de los trabajos se han llevado a cabo en los Estados Unidos de Norte América (EUA) con líneas o variedades adaptadas a las regiones templadas y el número de poblaciones de teocintle ha sido limitado. En México, hay poca información sobre los sistemas de incompatibilidad; el único trabajo que incluye datos de un número amplio de razas y accesiones de maíz de México fue llevado a cabo por Cíntora (1963), quien utilizó más de 200 accesio-nes del programa interamericano de maíz. Durante la última década no se han publi-cado estudios que involucren poblaciones de maíz y teocintle.
La presente investigación se llevó a cabo con la finalidad de determinar la constitución genética de 60 poblaciones de teocintle de México, respecto al Factor Gametofítico-1 (ga1).
Materiales y método
LOS SITIOS DE ESTUDIO:Los experimentos se realizaron en el Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA) de la Universidad de Guadalajara (UDG) en las Agujas, Nextipac, municipio de Zapopan, Jalisco y en el Centro de Investigaciones de la empresa Pioneer Hi-Bred International en Tapachula, Nayarit, México. El CUCBA

El factor gametofítico-1 (ga1) en teocintle (Zea spp.) en México 131
scientia-CUCBA 9(2):129—141.2007
se localiza a 20° 45’ N y 103° 31’ O, a una altitud de 1650 m, la temperatura media durante junio a octubre es de 23.6 °C y la cantidad de lluvia media anual es de 816 milímetros. Tapachula está situada a 20° 49’ N y 105° 55’ O, a una altitud de 26 m; la temperatura media, durante el periodo de enero a junio es de 25 °C y por lo ge-neral no se registran lluvias en ese lapso. Las siembras fueron establecidas en los ciclos Primavera-Verano de 2003, 2004 y 2005 en el CUCBA y en el ciclo de Otoño-Invierno 2004 en Tapachula.
MATERIAL GENÉTICO
Se usaron probadores desarrollados con líneas de germoplasma tropical origina-rios de la Universidad de Carolina del Norte, Raleigh, EUA y líneas de origen templado provenientes de la Universidad de Wisconsin, Madison, EUA. La lista de los materiales usados como probadores se presenta en el cuadro 1, que incluye el tipo de material, composición alélica y origen. Los materiales genéticos de interés para determinar la constitución genotípica, res-pecto a incompatibilidad fueron 60 colec-tas de teocintle, provenientes de diferentes regiones de México.
Cuadro 1. Materiales utilizados como probado-res para incompatibilidad, incluyendo su tipo y composición alélica y origen.
Material Tipo Genotipo Origen2
NC348 Línea Ga1-S/Ga1-S NCSU
NC354 Línea Ga1-S /Ga1-S NCSU
W22-P4830 Línea Ga1-S /Ga1-S WIS
W22-P5270 Línea Ga1-S /Ga1-S WIS
W22 Línea ga1/ga1 WIS
P-3394 Híbrido ga1/ga1 Pioneer
JAL-304 (Maíz Dulce)
Raza Ga1-S/Ga1-S CIMMYT
CHIH-147 (Azul)
Raza ga1/ga1 CIMMYT
2NCSU: North Carolina State University, Raleigh, USA.WIS: Universidad de Wisconsin, Madison, USA. CIMMYT: Centro Internacional de Mejoramiento de Maíz y Trigo.Pioneer Hi Bred International, USA.
IDENTIFICACIÓN DE ALELOS EN LAS POBLACIONES DE TEOCINTLE:La caracterización alélica de las poblacio-nes de teocintle se llevaron a cabo durante los ciclos de Otoño-Invierno de 2004 en Tapachula y en el Verano de 2004 y 2005 en el CUCBA utilizando probadores macho y hembra.
(i) Como probador macho se usó el híbri-do P-3394 (ga1/ga1), que proporcionó el polen a cada uno de los materiales de interés, para separar plantas con los alelos Ga1-m o ga1 (con producción de grano) de las plantas con el alelo Ga1-S (sin formación de grano).
(ii) Como probadores hembra (Ga1-S/Ga1-S), se usaron las cruzas NC354 × W22-P4830 y NC354 × W22-P5270, que recibieron polen de cada uno de los materiales de interés, para separar las plantas con el alelo Ga1-m (con llenado de mazorca) de aquellas con el alelo ga1 (sin llenado de mazorca).
Las cruzas con el probador macho se realizaron durante 2004 en un lote aislado en el campo experimental de la empresa Pioneer en Tapachula, Nay. Se sembraron 92 colectas de teocintle en parcelas de un surco de 5 m en bloques de cuatro surcos “hembra” para ser utilizados como pro-genitores femeninos, y cuatro surcos con tres fechas de siembra (a tiempo, ocho y 16 días después de la siembra de la hembra) del probador macho P-3394; previamente a la liberación del polen, las espigas de las plantas en cada uno de los surcos hembra fueron eliminadas. Simultáneamente, se sembró un lote de polinización libre con las mismas 92 colectas de teocintle para usarse como referencia en la evaluación del llenado de grano.
En el caso de las cruzas con los proba-dores hembra en el CUCBA durante 2004 y 2005, se sembraron 158 poblaciones de teocintle en dos surcos de 5 m, mientras que cada probador se sembró en 20 surcos de 5 m en cuatro fechas de siembra para

132 Lino De la Cruz Larios et al.
scientia-CUCBA 9(2):129—141.2007
asegurar buena sincronía en floración (a tiempo, 8, 15 y 22 días después a la siem-bra). Se hicieron cruzamientos manuales planta a planta colectando polen de cinco plantas de las poblaciones de teocintle para llevar a cinco plantas de cada uno de los probadores hembra NC354 × W22-P4830 y NC354 × W22-P5270. Por lo general, las polinizaciones manuales se llevaron a cabo entre las 12:00 y 14:00 horas en estigmas de 5 a 8 cm de longitud.
EVALUACIÓN DEL LLENADO DE MAZORCA EN MAÍZ
La formación de grano fue el dato clave para definir la constitución alélica a través de las cruzas con los probadores macho y hembra. Cada una de las mazorcas pro-veniente de los cruzamientos con los dos probadores hembra fue examinada indi-vidualmente. La estimación de llenado de mazorca se llevó a cabo visualmente re-dondeando con aproximación al 10% tal y como lo describió Kermicle y Allen (1990). Esto es, llenado abajo del 5% fue asignado 0%, de 5 a 15% el valor fue 10% y así en adelante hasta avanzar al 95% el cual fue asignado 100%. Se tomó como base el lado de la mazorca con el mejor llenado en toda su longitud. Adicionalmente a la estima-ción de llenado de mazorca, se tomaron fotografías de las mazorcas individuales resultantes de las cruzas de los materiales de interés con los tres probadores.
ESTIMACIÓN DE LA FORMACIÓN DE GRANO E IDENTIFICACIÓN DE ALELOS EN TEOCINTLE
Existen varias diferencias morfológicas entre el maíz y el teocintle por las que se requiere un manejo especial de los lotes aislados o polinizaciones manuales. El teocintle tiene ramas laterales que normal-mente terminan en espigas masculinas; en los nudos de cada rama lateral se desarro-llan inflorescencias femeninas con una o varias mazorcas (figura 1). En función de la competencia entre plantas y condiciones de manejo puede haber hasta 10 ramas laterales, 20 hijos y hasta cerca de 100 nu-dos con mazorca. Puede haber hasta cinco
mazorcas por nudo y cada mazorca con-tiene 6 a 12 semillas (Sánchez et al. 1998); todas estas características dificultan tanto la eliminación de espigas en lotes aislados así como cubrir las mazorcas y realizar polinizaciones controladas. En teocintle la semilla es una cariopsis simple encerra-da en una cápsula dura consistente de un segmento del raquis y una gluma inferior fuertemente endurecida; en maíz existe la copilla y las glumas, pero son de menor tamaño y no cubren el grano. Además, a lo descrito arriba, existen poblaciones prove-nientes de zonas cálidas de baja altitud con ciclo vegetativo muy largo con las cuales no se logran coincidencias en floración con facilidad.
Durante el invierno de 2004 en Tapachula, Nayarit, se sembraron 92 ac-cesiones de teocintle en un lote de polini-zación libre, se cosecharon únicamente 72 accesiones, para estimar el porcentaje de formación de grano, se calculó el índice (peso/volumen)×100 de la semilla produci-da por parcela. Con la finalidad de estimar el porcentaje de formación de grano, las semillas obscuras o negras corresponden a semillas completas, mientras que las semi-llas blancas están vacías (vanas). Con base en un clasificador de semillas (Seedburo Riffle Divider Mod. R75), se obtuvieron cuatro muestras al azar de 300 a 400 semi-llas por parcela. De cada una de las mues-tras, se contó el número total de semillas, blancas y negras.
ANÁLISIS ESTADÍSTICOS
Los datos de llenado de mazorca con cada probador hembra fueron analizados con base en dos procedimientos:
(i) Se llevó a cabo un análisis de varianza con la fuente de variación de teocintle y las medias se compararon con base a la prueba de Dunnett al 0.05 de proba-bilidad, usando el Procedimiento GLM del Sistema de Análisis Estadístico (SAS Institute 1992); la prueba se llevó a cabo con respecto a cruzas de prue-ba tanto para ga1 como para Ga1-S

El factor gametofítico-1 (ga1) en teocintle (Zea spp.) en México 133
scientia-CUCBA 9(2):129—141.2007
Figura 1. Lote de desespigamiento para estimación de la formación de grano en teocintle.

134 Lino De la Cruz Larios et al.
scientia-CUCBA 9(2):129—141.2007
(JAL-304 Ga1-S/Ga1-S y CHIH-147 ga1/ga1). Además, se hicieron compa-raciones entre proporciones de llenado con base en una prueba aproximada de Z (Hicks 1982).
(ii) Las colectas de teocintle fueron agru-padas con base en sus relaciones genéticas y origen geográfico respecti-vamente. Las diferencias entre los gru-pos para la variable llenado de mazor-ca fueron determinadas con base en la diferencia mínima significativa (DMS) usando los procedimientos GLM y MIXED del SAS.
REGRESIÓN
Los análisis estadísticos de los datos del teocintle fueron orientados para determi-nar la relación entre el índice de peso/vo-lumen y la formación de grano; dicha rela-ción se determinó por medio de regresión lineal simple del porcentaje de semillas negras en el índice de peso/volumen del lote aislado. La línea de regresión se cal-culó con base en los datos de 72 poblacio-nes que tuvieron una buena sincronía en floración con el probador macho P-3394. La estimación de la formación de grano en teocintle representa la primera fase en el entendimiento de la compatibilidad. Los resultados en teocintle pueden ser inter-pretados en forma preliminar como sigue: 1. Si se observa que el progenitor femeni-
no (teocintle) produce semilla completa
(negra) existen dos posibilidades de ale-los: el progenitor femenino es recesivo ga1 o Ga1-m. Cruzando el teocintle con un probador femenino Ga1-S podríamos distinguir ga1 de Ga1-m.
2. Si no se observa ninguna semilla en el progenitor femenino (teocintle) o la se-milla es blanca, entonces su genotipo es Ga1-S/Ga1-S
3. Cuando se tiene llenado de grano par-cial, las interpretaciones se complican dado que los alelos que tienen actividad vía polen (Ga1-m) o Ga1-S podrían estar involucrados.
Resultados
ESTIMACIÓN DE LLENADO DE GRANO EN TEOCINTLE
En el cuadro 2 se presentan las medias y rangos para la formación de grano en teo-cintle, obtenidos en un lote de polinización libre y en un lote aislado en Tapachula, Nay. durante el ciclo de invierno de 2004. En la figura 1 se presenta una panorámica del lote de desespigamiento del teocintle en Tapachula, Nayarit.
Los rangos de formación de grano son muy amplios para las razas Mesa Central (ssp. mexicana) y Balsas (ssp. par-viglumis); los valores más bajos en la raza Mesa Central fueron para poblaciones cer-canas al lago de Cuitzeo. En la raza Balsas se encontró un rango similar al de Mesa
Cuadro 2. Estimación de formación de grano en teocintle. Tapachula, Nayarit. 2004.
Raza Número colectas
Índice lote aislado Índice* P.L. % Grano negro
Media Rango Media Rango
Chalco 17 21.3 14.8 - 32.0 63.8 13.6 3.5 - 29.2
Mesa Central 24 30.3 16.5 - 66.2 69.4 24.1 5.5 - 82.5
Durango 3 56.7 54.7 - 59.2 71.4 59.3 52.2 - 66.1
Nabogame 2 40.1 38.0 - 42.1 57.7 43.2 40.8 - 45.5
Balsas 14 43.0 25.1 - 62.5 73.5 42.0 18.2 - 83.6
*Indice en el lote de polinización libre (P.L.)
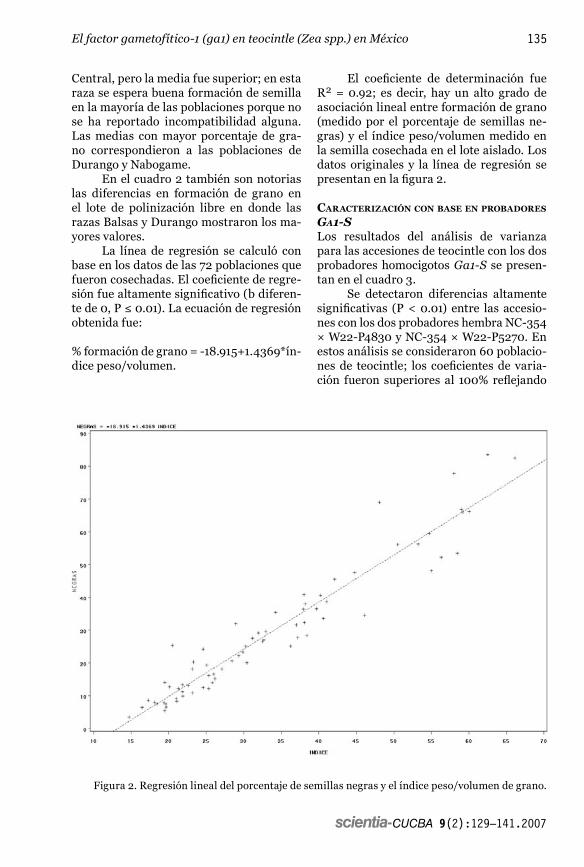
El factor gametofítico-1 (ga1) en teocintle (Zea spp.) en México 135
scientia-CUCBA 9(2):129—141.2007
Central, pero la media fue superior; en esta raza se espera buena formación de semilla en la mayoría de las poblaciones porque no se ha reportado incompatibilidad alguna. Las medias con mayor porcentaje de gra-no correspondieron a las poblaciones de Durango y Nabogame.
En el cuadro 2 también son notorias las diferencias en formación de grano en el lote de polinización libre en donde las razas Balsas y Durango mostraron los ma-yores valores.
La línea de regresión se calculó con base en los datos de las 72 poblaciones que fueron cosechadas. El coeficiente de regre-sión fue altamente significativo (b diferen-te de 0, P ≤ 0.01). La ecuación de regresión obtenida fue:
% formación de grano = -18.915+1.4369*ín-dice peso/volumen.
El coeficiente de determinación fue R2 = 0.92; es decir, hay un alto grado de asociación lineal entre formación de grano (medido por el porcentaje de semillas ne-gras) y el índice peso/volumen medido en la semilla cosechada en el lote aislado. Los datos originales y la línea de regresión se presentan en la figura 2.
CARACTERIZACIÓN CON BASE EN PROBADORES GA1-SLos resultados del análisis de varianza para las accesiones de teocintle con los dos probadores homocigotos Ga1-S se presen-tan en el cuadro 3.
Se detectaron diferencias altamente significativas (P < 0.01) entre las accesio-nes con los dos probadores hembra NC-354 × W22-P4830 y NC-354 × W22-P5270. En estos análisis se consideraron 60 poblacio-nes de teocintle; los coeficientes de varia-ción fueron superiores al 100% reflejando
Figura 2. Regresión lineal del porcentaje de semillas negras y el índice peso/volumen de grano.

136 Lino De la Cruz Larios et al.
scientia-CUCBA 9(2):129—141.2007
la gran diversidad genética entre y dentro de razas y poblaciones de teocintle para las reacciones de incompatibilidad.
En el cuadro 4 se presentan las me-dias de formación de grano con los tres probadores. Los análisis con los probado-res hembra Ga1-S/Ga1-S mostraron que el 23% de las muestras evaluadas tuvieron un llenado significativamente igual al llenado con el testigo JAL-304 (Ga1-S/Ga1-S); la mayor parte de esas accesiones tuvieron porcentajes bajos de formación de grano cuando se usaron como hembras con P-3394. De las 60 accesiones en que se tuvo información con todos los probadores, en 34 se detectó el alelo Ga1-S (57%), de las cuales 14 (23%) fueron clasificadas como homocigotas Ga1-S/Ga1-S. En 68% de las accesiones se detectó el alelo ga1, pero úni-camente 11 (18%) fueron homocigotas. Por último, 15 de las accesiones presentaron el alelo Ga1-m (25%), de las cuales 5 (8%) fueron homocigotas Ga1-m/Ga1-m.
DIFERENCIAS ENTRE RAZAS
Las accesiones representativas de teocintle de México se dividieron en razas y se lleva-ron a cabo los análisis de varianza con dos probadores hembra. Se detectaron diferen-cias altamente significativas entre razas y entre accesiones dentro de razas para los dos probadores (cuadro 5). Los coeficientes de variación fueron altos para los datos de
Cuadro 3. Cuadrados medios del análisis de varianza y valores de Dunnett (5%) para forma-ción de grano (%) de teocintle con los probado-res hembra NC-354 × W22-P4830 y NC-354 × W22-P5270.
F.V. G.L. P4830 P5270
(Ga1-S/Ga1-S)
Teocintle 59 2495.1** 3568.23**
Error 423 679.45 1159.24
C.V.(%) 136.4 134.11
Dunnett (5%) 3.07 2.99
** Altamente significativos (P < 0.01)
llenado de mazorca con los dos probadores hembra P4830 y P5270.
Las medias para la formación de grano para los dos probadores se presen-tan para cada raza y región en el cuadro 6. Las regiones de Guachinango-Amatlán de Cañas (Balsas) y Durango (Mesa Central) y Nabogame presentaron los porcentajes más bajos de llenado de mazorca con res-pecto al probador femenino P-4830 con 0.7 y 0.6 y 0% respectivamente, en estas re-giones predominan los genotipos ga1/ga1; para este mismo probador femenino, la región de Opopeo y Ciudad Hidalgo (Mesa Central) fue la que presentó el porcentaje más alto de llenado de mazorca con 56%
El llenado de mazorca con el pro-bador P-5270 fue de igual forma muy variable entre regiones destacando el porcentaje de llenado más alto en las re-giones de Tejupilco-Palmar Chico (Balsas), Puruándiro (Mesa Central) y Malinalco (Balsas); en estas regiones es posible en-contrar poblaciones con alelos de incom-patibilidad Ga1-S.
Discusión
Con base en los resultados anteriores, se puede decir que la distribución geográfica de los alelos del locus ga1 no es aleatoria y se pudieron observar patrones definidos en México. El alelo recesivo ga1 está presen-te en bajas proporciones en el Occidente de México en los estados de Jalisco y Michoacán y parece estar fijo en las po-blaciones de teocintle de los estados de Durango y Chihuahua. Por su parte el alelo Ga1-S está distribuido casi exclusivamente en las razas Chalco y Mesa Central en la región centro del país (Estado de México, Puebla, Michoacán, Guanajuato y Jalisco).
La distribución para el alelo Ga1-m es más limitada al encontrarse en muy po-cas accesiones sólo en el Estado de México, Michoacán y Jalisco.
El alelo Ga1-S responsable de la in-compatibilidad no recíproca estuvo presen-te en casi todas las poblaciones de teocintle tipo maleza (ssp. mexicana); por su parte
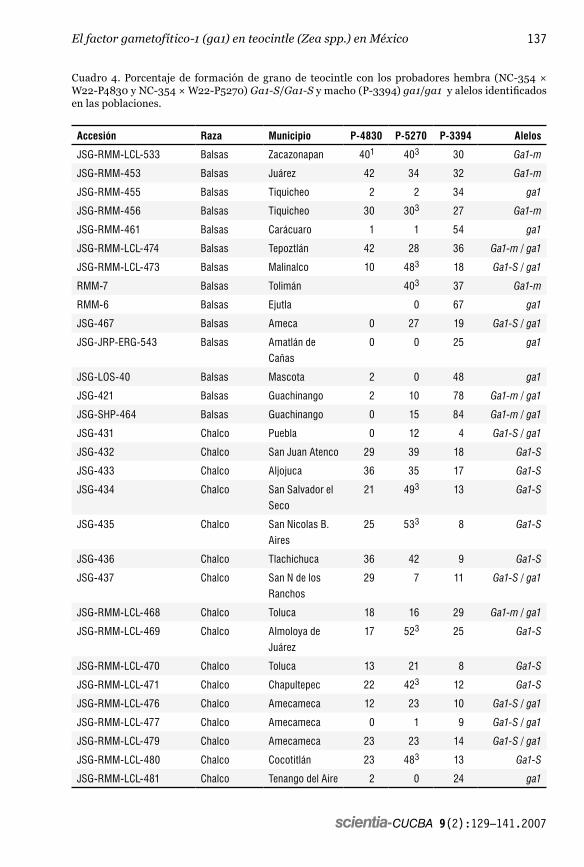
El factor gametofítico-1 (ga1) en teocintle (Zea spp.) en México 137
scientia-CUCBA 9(2):129—141.2007
Cuadro 4. Porcentaje de formación de grano de teocintle con los probadores hembra (NC-354 × W22-P4830 y NC-354 × W22-P5270) Ga1-S/Ga1-S y macho (P-3394) ga1/ga1 y alelos identificados en las poblaciones.
Accesión Raza Municipio P-4830 P-5270 P-3394 Alelos
JSG-RMM-LCL-533 Balsas Zacazonapan 401 403 30 Ga1-m
JSG-RMM-453 Balsas Juárez 42 34 32 Ga1-m
JSG-RMM-455 Balsas Tiquicheo 2 2 34 ga1
JSG-RMM-456 Balsas Tiquicheo 30 303 27 Ga1-m
JSG-RMM-461 Balsas Carácuaro 1 1 54 ga1
JSG-RMM-LCL-474 Balsas Tepoztlán 42 28 36 Ga1-m / ga1
JSG-RMM-LCL-473 Balsas Malinalco 10 483 18 Ga1-S / ga1
RMM-7 Balsas Tolimán 403 37 Ga1-m
RMM-6 Balsas Ejutla 0 67 ga1
JSG-467 Balsas Ameca 0 27 19 Ga1-S / ga1
JSG-JRP-ERG-543 Balsas Amatlán de
Cañas
0 0 25 ga1
JSG-LOS-40 Balsas Mascota 2 0 48 ga1
JSG-421 Balsas Guachinango 2 10 78 Ga1-m / ga1
JSG-SHP-464 Balsas Guachinango 0 15 84 Ga1-m / ga1
JSG-431 Chalco Puebla 0 12 4 Ga1-S / ga1
JSG-432 Chalco San Juan Atenco 29 39 18 Ga1-S
JSG-433 Chalco Aljojuca 36 35 17 Ga1-S
JSG-434 Chalco San Salvador el
Seco
21 493 13 Ga1-S
JSG-435 Chalco San Nicolas B.
Aires
25 533 8 Ga1-S
JSG-436 Chalco Tlachichuca 36 42 9 Ga1-S
JSG-437 Chalco San N de los
Ranchos
29 7 11 Ga1-S / ga1
JSG-RMM-LCL-468 Chalco Toluca 18 16 29 Ga1-m / ga1
JSG-RMM-LCL-469 Chalco Almoloya de
Juárez
17 523 25 Ga1-S
JSG-RMM-LCL-470 Chalco Toluca 13 21 8 Ga1-S
JSG-RMM-LCL-471 Chalco Chapultepec 22 423 12 Ga1-S
JSG-RMM-LCL-476 Chalco Amecameca 12 23 10 Ga1-S / ga1
JSG-RMM-LCL-477 Chalco Amecameca 0 1 9 Ga1-S / ga1
JSG-RMM-LCL-479 Chalco Amecameca 23 23 14 Ga1-S / ga1
JSG-RMM-LCL-480 Chalco Cocotitlán 23 483 13 Ga1-S
JSG-RMM-LCL-481 Chalco Tenango del Aire 2 0 24 ga1

138 Lino De la Cruz Larios et al.
scientia-CUCBA 9(2):129—141.2007
Cuadro 4. Continuación.
Accesión Raza Municipio P-4830 P-5270 P-3394 Alelos
JSG-RMM-LCL-482 Chalco Temamatla 0 14 8 Ga1-S / ga1
JSG-427 Mesa Central Ixtlán 27 25 28 Ga1-S / ga1
JSG-465 Mesa Central Ayotlán 0 0 82 ga1
JSG-MAS-ERV-411 Mesa Central La Barca 24 403 12 Ga1-S
JSG-ERV-409 Mesa Central Poncitlán 20 533 22 Ga1-S / ga1
JSG-426 Mesa Central Churintzio 27 20 32 Ga1-m
JSG-RMM-438 Mesa Central Churintzio 34 28 21 Ga1-S / ga1
JSG-RMM-439 Mesa Central Churintzio 4 27 39 Ga1-m / ga1
JSG-RMM-440 Mesa Central Penjamillo 9 40 20 Ga1-S / ga1
JSG-RMM-442 Mesa Central Zináparo 2 35 32 Ga1-m / ga1
JSG-RMM-452 Mesa Central Churintzio 26 22 28 Ga1-m / ga1
JSG-RMM-441 Mesa Central Puruándiro 41 523 8 Ga1-S
JSG-RMM-443 Mesa Central Puruándiro 25 643,4 23 Ga1-S / ga1
JSG-RMM-444 Mesa Central Puruándiro 40 30 6 Ga1-S / ga1
JSG-RMM-445 Mesa Central Moroleón 0 24 13 Ga1-S / ga1
JSG-RMM-446 Mesa Central Moroleón 3 3 15 ga1
JSG-RMM-447 Mesa Central Uriangato 21 403 14 Ga1-S / ga1
JSG-RMM-448 Mesa Central Cuitzeo 1 17 20 Ga1-S / ga1
JSG-RMM-449 Mesa Central Huandacareo 511 12 16 Ga1-S / ga1
JSG-RMM-450 Mesa Central Copándaro 21 9 5 Ga1-S / ga1
JSG-RMM-451 Mesa Central Chucándiro 1 16 20 Ga1-S
JSG-RMM-LCL-536 Mesa Central Hidalgo 751,2 25 13 Ga1-S
JSG-RMM-463 Mesa Central Salvador
Escalante
37 533 11 Ga1-S
JSG-RMM-428 Mesa Central Durango 1 11 60 Ga1-m / ga1
JSG-RMM-429 Mesa Central Durango 1 20 66 Ga1-m / ga1
JSG-RMM-430 Mesa Central Durango 0 12 52 Ga1-m / ga1
JSG-ERV-407 Mesa Central La Barca 30 17 25 Ga1-S / ga1
JSG-MAS-ERV-412 Mesa Central Poncitlán 0 0 66 ga1
JSG-414 Nobogame Guadalupe y
Calvo
0 0 41 ga1
JSG-415 Nobogame Guadalupe y
Calvo
0 0 46 ga1
1 Promedio estadísticamente igual a la cruza testigo de JAL-304 por P4830 (Ga1-S/Ga1-S).2 Promedio estadísticamente diferente a la cruza testigo de CHIH-147 por P4830 (Ga1-S/Ga1-S).3 Promedio estadísticamente igual a la cruza testigo de JAL-304 con (P5270) Ga1-S/Ga1-S.4 Promedio estadísticamente diferente a la cruza testigo de CHIH-147 con (P5270) Ga1-S/Ga1-S.

El factor gametofítico-1 (ga1) en teocintle (Zea spp.) en México 139
scientia-CUCBA 9(2):129—141.2007
Cuadro 5. Cuadrados medios del análisis de va-rianza para regiones de teocintle.
F.V. G.L. P4830 P5270
(Ga1-S/Ga1-S)
Razas 4 3322.19** 4060.59**
Accesiones
/Razas
55 1985.46** 3157.62**
Error 644 689.23 1170.04
C.V. (%) 141.2 136.24
** Altamente significativos (P < 0.01)
el alelo Ga1-m se presentó únicamente en cerca del 25% de las accesiones de teocin-tle. La distribución más limitada del alelo Ga1-m y mayor frecuencia de Ga1-S son de importancia dado que puede tener relación con el aislamiento reproductivo entre maíz y teocintle. Estudios recientes indican que el polen Ga1-S fertiliza con más efi-ciencia óvulos Ga1-S/Ga1-S que el polen Ga1-m cuando los dos tipos de polen están presentes sobre los estigmas (Kermicle 2006). De cualquier manera, es necesario acumular más evidencias para asegurar que el locus ga1, en especial el alelo Ga1-S desempeña un papel determinante en ex-plicar el aislamiento reproductivo en maíz
Cuadro 6. Medias de formación de grano (%) por regiones con probadores hembra Ga1-S/Ga1-S (NC354 × W22-P4830 y NC354 × W22-P5270) y macho ga1/ga1 (P-3394).
Raza/Región Accesiones P4830 P5270 P3394
Balsas 14 14.2 19.5 42.0
Sur de Jalisco 2 20.0 51.7
Guachinango-A. de Cañas 5 0.7 10.3 50.8
Tejupilco-Palmar Chico 1 40.0 40.0 29.7
Huetamo-Carácuaro 4 18.5 16.7 36.4
Malinalco 1 10.0 47.7 18.2
Amatlán-Tepoztlán 1 42.3 27.8 36.5
Chalco 17 18.1 28.1 13.6
Valle de Toluca 4 17.7 32.8 18.6
Chalco-Amecameca 6 10.0 18.4 13.0
Puebla 7 25.1 33.7 11.3
Durango 3 0.6 14.1 59.3
Mesa Central 24 21.6 27.1 24.1
Cuitzeo 7 13.9 17.3 14.0
Churintzio-La Barca 12 17.0 25.5 34.0
Puruándiro 3 35.2 48.6 12.7
Opopeo-Cd. Hidalgo 2 56.0 39.0 12.2
Nobogame 2 0.0 0.0 43.2

140 Lino De la Cruz Larios et al.
scientia-CUCBA 9(2):129—141.2007
y teocintle, tal y como lo señalan Kermicle et al. (2006).
Por otra parte, es importante señalar que a diferencia de los estudios en maíz, las cruzas con los probadores cuando se usa como macho el teocintle mostraron valores bajos de llenado de mazorca; aun cuando se considera que están presentes en el po-len los alelos Ga1-m y Ga1-S que aseguran compatibilidad con cualquier tipo de estig-ma. Por una parte, el polen del teocintle es afectado en mayor grado que el maíz por la desecación, es de menor tamaño, y su tubo polínico tiene menor longitud que el polen del maíz (Rodríguez et al. 2006). De acuer-do con Aguirre (1977), en la primera etapa de desarrollo de los tubos polínicos, la ve-locidad de crecimiento es similar tanto en maíz como en teocintle, y esta velocidad va diferenciándose a medida que los tubos po-línicos avanzan en los estilos, habiendo un retraso en los tubos de teocintle. El retraso de este tubo polínico posiblemente sea cau-sado por cierto agotamiento de las reservas del polen de teocintle almacenadas en su citoplasma, ya que, como se señaló antes, el tamaño del polen de teocintle es menor que el del maíz. Por tanto, la poca capaci-dad del polen del teocintle para fecundar al maíz podría ser considerada una de las barreras activas de aislamiento reproduc-tivo que permite la infiltración genética entre las dos especies. Por lo anterior, es necesario intensificar los trabajos que per-mitan entender la interacción entre polen y pistilo, especialmente cuando se usa como progenitor masculino el teocintle y el maíz como progenitor femenino; es probable que se requiera desarrollar probadores de maíz con características especiales de estigmas y evitar cualquier interacción no deseada.
Conclusiones
Debido a las dificultades para hacer esti-maciones directas de formación de grano en teocintle, en el presente estudio se de-terminó que la relación de peso sobre vo-lumen de grano es un estimador altamente
correlacionado al porcentaje de llenado de grano.
La constitución alélica de las pobla-ciones de teocintle en México es muy va-riable; sin embargo, el alelo Ga1-S respon-sable de la incompatibilidad no-recíproca fue el más frecuente, predominando en las razas Chalco y Mesa Central.
En las razas Balsas, Durango y Nobogame se encontraron predominante-mente los alelos Ga1-m y el recesivo ga1.
En áreas de México donde se distri-buyen las especies silvestres con el alelo Ga1-S debería programarse la siembra de híbridos comerciales, usando exclusi-vamente los recesivos para el locus ga1. En los casos de poblaciones con los alelos Ga1-m y ga1, debería evitarse la siembra cercana de materiales mejorados introdu-cidos de otras áreas para minimizar flujo genético no deseado.
Bibliografía Aguirre G., C. D. 1977. Competencia entre el
polen de maíz y de teocintle durante la fe-cundación. Tesis de Maestría. Colegio de Postgraduados. Chapingo, México. 99 pp.
Cíntora O., C. 1963. Incompatibilidad no re-cíproca en algunas colecciones Mexicanas de maíz. Tesis Maestría Rama de Genética. Facultad del Colegio de Graduados de la E.N.A.Chapingo, México. 39 pp.
Doebley, J. F. 2004. The genetics of maize evolution. Annual Review of Genetics 38: 37–59.
Evans, M. M. S. y J. L. Kermicle. 2001. Teosinte crossing barrier1, a locus gover-ning hybridization of teosinte with maize. Theoretical and Applied Genetics 103: 259–265.
Hicks, C. R. 1982. Fundamental concepts in the design of experiments. Holt, Rinehart and Winston, Inc, New York. 425 pp.
Kermicle, J. L y J. O. Allen. 1990. Cross-in-compatibility between maize and teosinte. Maydica 35: 399–408.
Kermicle, J. L. 2006. A selfish gene governing pollen-pistil compatibility confers repro-ductive isolation between maize relatives. Genetics 172: 499–506.
Kermicle, J. L., S. Taba y M. M. S. Evans. 2006. The GAMETOPHYTE-1 locus and re-

El factor gametofítico-1 (ga1) en teocintle (Zea spp.) en México 141
scientia-CUCBA 9(2):129—141.2007
productive isolation among Zea mays subs-pecies. Maydica 51: 219–226.
Mangelsdorf, P. C., R. S. MacNeish y W. C. Galinat. 1967. Prehistoric maize, Teocintle and Tripsacum from Tamaulipas, Mexico. Botanical Museum Leaflets Harvard University 22: 33–63.
Matsuoka, Y., Y. Vigouroux, M. M. Goodman, J. J. Sánchez G., E. Bukler y J. F. Doebley. 2002. A single domestication for maize shown by multilocus microsatelli-te genotyping. Proceedings of the National Academy of Sciences 99: 6080–6084.
Nelson, O. E. 1996. The gametophyte factors of maize. In: M. Freeling & V. Walbot (eds.) The Maize Handbook. Springer-Verlag. pp. 496–503.
Rodríguez F., J. G., J. J. Sánchez G., B. Baltazar M., L. de la Cruz L., F. Santacruz R., J. Ron P. y J. B. Schoper. 2006. Characterization of floral morpho-logy and synchrony among Zea species in Mexico. Maydica 51: 383–398.
SAS Institute, Inc. 1992. SAS Technical Report P-229. SAS/STAT Software: Changes and Enhancements. SAS Institute, Inc., Cary, NC.
Sánchez G., J. J., T. A. Kato Y., M. Aguilar S., J. M. Hernández C., A. López R y J. A. Ruíz C. 1998. Distribución y carac-terización del teocintle. Libro Técnico No. 2. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). 149 pp.
Wilkes, H. G. 1977 Hybridization of maize and teosinte, in Mexico and Guatemala and the improvement of maize. Economic Botany 31: 254–293.
FECHA EFECTIVA DE PUBLICACIÓN 29 DE DICIEMBRE DE 2007

142
scientia-CUCBA 9(2):142.2007

scientia-CUCBA 9(2):143—152.2007
Efecto de la aplicación de compostas sobre las características físicas, químicas y biológicas del suelo en el cultivo de frijol
Martha Delia Ocegueda Reyes1, Eduardo López Alcocer2, Rogelio Lépiz Ildefonso3, Diego R. González Eguiarte4, Ricardo Nuño Romero4, Fernando López Alcocer4 y Patricia Zarazúa Villaseñor4
1Posgrado en Ciencias Agrícolas y Forestales, CUCBA. Las Agujas, Zapopan, Jal. 2Departamento de Ciencias Ambientales, CUCBA. 3Departamento de Producción Agrícola, CUCBA. 4Departamento de Desarrollo Rural Sustentable, CUCBA. Las Agujas, Municipio de Zapopan, Jal. Correo electrónico: [email protected], [email protected], [email protected], [email protected]
Resumen: El presente trabajo se realizó con los objetivos de determinar el efecto de la aplicación de cuatro compostas producidas a partir de diferente materia prima, sobre algunas características físicas, químicas y biológicas del suelo y sobre el crecimiento de plantas de frijol, bajo condiciones de invernadero. Los tratamientos fueron: (1) composta tipo Bocashi; (2) composta a base de paja de frijol; (3) de paja de frijol-maíz; (4) paja de maíz; (5) vermiabono; (6) fertilización química y (7) testigo. En el suelo se evaluaron variables físicas, químicas y microbiológicas, así como algunas variables en la planta de frijol. Los resultados mostraron que la aplicación de compostas al suelo presentan una tendencia a favorecer la capacidad de absorción de agua y a disminuir la densidad aparente. No se encontraron diferencias significativas de la aplicación de compostas sobre las variables químicas de suelo. En las variables microbiológicas se encontraron diferencias estadísticas significativas para las poblaciones de bacterias y actinomicetos, no así para la población de hongos. En las variables evaluadas en la planta de frijol, se encontraron diferencias significativas en la nodulación y rendimiento de grano. Los resultados sugieren que para inducir cambios en las características físicas, químicas y biológicas del suelo, se requiere de la aplicación periódica de mayores volúmenes de composta. Se encontró, además, que la aplicación de abonos orgánicos puede disminuir o sustituir la aplicación de fertilizante químico en la producción de grano en el cultivo de frijol.
Palabras clave: Frijol, composta y población microbiana.
Abstract: The present study was carried out with a view to determine the effect of the application of four composts produced from different raw material, on some physical, chemical and biological characteristics of the soil and on the growth of bean plants, under greenhouse conditions. The treatments were: (1) compost Bocashi type; (2) compost with bean straw; (3) bean-maize straw; (4) maize straw; (5) casting; (6) chemical fertilization and (7) only soil (control). Physical, chemical and microbiological characteristics of the soil, as well as some variables in the bean plant, were evaluated. The results showed that the application of compost to the ground presents a tendency to favor the capacity of water absorption and to diminish the apparent density. Significant differences of the application of compost were not in the chemical soil variables. In the microbiological variables, there were significant statistical differences for the populations of bacteria and actinomycetes; not so for the population of fungi. In the variables evaluated in the bean plant, there were significant differences in the nodulation and grain yield. The results suggest that to cause a greater effect on the physical, chemical and biological soil properties, the application of greater volumes of compost is required. Also yielded from

144 Martha Delia Ocegueda Reyes et al.
scientia-CUCBA 9(2):143—152.2007
the study is evidence that the application of organic matter can reduce or replace the application of chemical fertilizer for the production of grain in bean crop.
Key words: Bean, compost and microbial population.
Introducción
En los sistemas de producción agrícola, en las últimas cinco décadas, se ha gene-ralizado el uso intensivo de agroquímicos (fertilizantes químicos, herbicidas, insecti-cidas y fungicidas). Como consecuencia de ello, a partir de la década de los 70’s em-pezaron a manifestarse fuertes problemas ambientales por la degradación de los re-cursos naturales (López et al 2001; López et al. 2004). En el suelo, la degradación se manifiesta por la pérdida de la vegetación nativa, altos niveles de erosión, disminu-ción de la disponibilidad de humedad, dis-minución de la materia orgánica y la acti-vidad microbiana, así como el incremento de acidez, factores que provocan la pérdida de la fertilidad del suelo (Alexander 1977; Velásquez 1997).
Una opción para rehabilitar los suelos afectados por los problemas de degrada-ción, es la incorporación de materia orgá-nica (Labrador 1996; Calderón et al. 2004; López et al. 2004), mediante la aplicación de abonos orgánicos, ya sea en forma de estiércoles, abonos verdes, composta, ver-miabono u otros; de esta manera, además de aportar nutrimentos para el desarrollo y producción de los cultivos, se mejoran las condiciones físicas de los suelos (Labrador 1996; Avilés y Tello 2001).
La incorporación de materiales orgánicos al suelo ha sido una práctica común para mejorar su fertilidad; así lo manifiestan Torres et al. (2002), quienes observaron la presencia de nódulos de bac-terias en la raíz del frijol y un incremento del nitrógeno al utilizar residuos orgánicos de sorgo (Sorghum bicolor L. Moench) y Crotalaria (Crotalaria juncea) marcados con 15Nitrógeno. Estos autores recomen-daron la incorporación de residuos de
sorgo para mejorar las condiciones físicas y químicas del suelo.
Por otra parte, la fertilidad del sue-lo está estrechamente relacionada con la población microbiana del mismo; también en este caso la incorporación de materia orgánica al suelo incrementa la población y actividad microbiana, produce benefi-cios adicionales a los relacionados con las características físicas y químicas (Triano et al. 2005). De acuerdo con Dalzell et al. (1991), las compostas contienen altas po-blaciones de microorganismos, por lo que uno de los objetivos de su aplicación al sue-lo es añadir vida microbiana y suministrar alimento para los microorganismos ya pre-sentes. Específicamente los hongos y acti-nomicetos actúan a través de la adherencia de sus hifas en las partículas del suelo, de esta forma mejoran los canales para el movimiento del aire, del agua y promueven un mayor crecimiento de las raíces de las plantas (Dalzell et al. 1991)
La materia orgánica influye en la for-mación de una buena estructura, en el me-joramiento del pH y en la capacidad de re-tención de agua, propiedades que facilitan el manejo y mejoran la productividad de los suelos (Kononova 1965; Luna y Vázquez 2003). También existen evidencias sobre la influencia de la materia orgánica en la reducción de problemas relacionados con plagas y enfermedades (Queitsch 2005).
Los efectos positivos de la aplicación de abonos orgánicos sobre el incremento en el rendimiento de grano de los cultivos, lo asientan López et al. (2001). Se informa que la aplicación de 2 t ha-1 de cachaza al momento de la siembra inducen la dismi-nución de la conductividad eléctrica (CE), el incremento de la materia orgánica, el in-cremento de elementos nutricionales como

Efecto de la aplicación de compostas sobre las características físicas... 145
scientia-CUCBA 9(2):143—152.2007
potasio, magnesio y nitrógeno, así como el incremento en rendimiento de grano en maíz (Arrieche y Mora 2005); también se consigna la disminución en la velocidad de infiltración del agua, una reducción en las pérdidas por evaporación y un incremento de la productividad en el cultivo del ciruelo (Eduardo et al. 2001).
Ante el problema de la degradación de los suelos en la región de Zapopan, se planteó el presente trabajo en el que se uti-lizó como indicador al cultivo de frijol, con el objetivo de determinar el efecto de la aplicación de diferentes compostas sobre algunas características físicas, químicas y biológicas del suelo y evaluar el efecto so-bre el desarrollo de la planta de frijol, bajo condiciones de invernadero.
Materiales y métodos
El presente trabajo se realizó en inver-nadero, así como en los laboratorios de Agromicrobiología y de Suelos del Centro Universitario de Ciencias Biológicas y Agropecuarias (CUCBA), de la Universidad de Guadalajara ubicado en Las Agujas, mu-nicipio de Zapopan, Jalisco, localizado en las coordenadas geográficas 20° 44́ Norte, 103° 30́ Oeste, y una altitud de 1570 m.
Se ensayaron siete tratamientos: 4 compostas elaboradas a partir de diferen-tes materiales, dos tratamientos a base de vermiabono y fertilizante químico y un
testigo (cuadro 1). Se utilizó un diseño ex-perimental completamente al azar con 10 repeticiones. Las unidades experimentales (UE) fueron bolsas de plástico negro de 20 × 35 cm, con 4.0 kg de capacidad. El suelo se tomó de una parcela vecina. Se sem-braron tres semillas de frijol por maceta, variedad Azufrado Tapatío, el 16 de agosto de 2006. Una vez establecido el cultivo, se conservó para evaluación una planta por maceta.
Para el suelo se registraron variables químicas, físicas y biológicas. Las variables químicas se especifican en el cuadro 2 y se determinaron según el método de Richards (1982). Las variables físicas consideradas fueron: capacidad de absorción de agua (g cm-3), capilaridad (g cm-3), densidad (g cm-3) y tiempo de infiltración (minutos), determinadas según el manual de Fisher et al. (2003). Las variables microbiológicas fueron: población de hongos, bacterias y actinomicetos, valores consignados como unidades formadoras de colonias (UFC); se utilizaron los medios de cultivo PDA, Agar nutritivo y Czapek, respectivamente. El método fue de diluciones decimales y vaciado en placa (Valdés et al. 2003).
En la planta de frijol se registraron siete variables, cinco en la etapa vegetati-va: longitud de parte área, medida desde el hipocotilo hasta el extremo superior de la última hoja trifoliada; longitud de raíz, medida desde el hipocotilo hasta el extre-
Cuadro 1. Tratamientos evaluados en el experimento de abonos orgánicos.
No. de tratamiento
Tratamiento Dosis
1 Composta tipo Bocashi, compuesta de Paja de maíz, paja de fríjol, estiércol,
levadura y melaza.
10 t ha-1
2 Composta a base de paja de fríjol y estiércol 10 t ha-1
3 Composta a base de paja de maíz, paja de frijol y estiércol 10 t ha-1
4 Composta a base de paja de maíz y estiércol 10 t ha-1
5 Vermiabono (abono de lombriz) 10 t ha-1
6 Fertilizante químico 60-60-00
7 Testigo -

146 Martha Delia Ocegueda Reyes et al.
scientia-CUCBA 9(2):143—152.2007
mo inferior de la raíz principal; peso de materia seca de la parte aérea de la planta (biomasa); peso seco de la raíz y número de nódulos por planta. En madurez de co-secha: número de vainas y peso de grano. En ambas etapas el número de plantas eva-luadas por tratamiento fue de cinco. Las observaciones de las variables de planta y microbiológicas, se sometieron a análisis de varianza y prueba de separación de me-dias: Tukey 0.05 (Steel y Torrie 1985).
Resultados y discusión
De las variables físicas y químicas tomadas del suelo, sólo la capacidad de absorción de agua y el N-NO3 mostraron diferencia sig-nificativa para tratamientos. Sin embargo, se presentan graficas de tendencia para las variables de mayor relevancia.
Cuadro 2. Variables químicas determinadas en el suelo.
Variable Método
Materia Orgánica (%) Walkey-Black
Capacidad de Intercambio
Catiónico (meq 100 g
suelo-1)
Acetato de amonio
Cationes intercambiables:
Ca (meq 100 g suelo-1) E.D.T.A.
Mg (meq 100 g suelo-1) Calculado
K (meq 100 g suelo-1) Flamometría
pH Potenciómetro
N nítrico (ppm) Morgan
N amoniacal (ppm) Morgan
P (ppm) Morgan
K (ppm) Morgan
Ca (ppm) Morgan
Mg (ppm) Morgan
Conductividad Eléctrica (CE)
(mmhos cm2 a 25 ºC)
Conductímetro
Relación C/N Calculado
La figura 1 muestra los resultados de la variable absorción de agua, donde los tratamientos 1 (composta tipo Bocashi) y 5 (vermiabono), mostraron los valores más altos, en tanto que al tratamiento 7 (Testigo) sin aplicación de abonos orgá-nicos, correspondió el valor más bajo. El resultado concuerda con la literatura, en el sentido de que la materia orgánica favore-ce la capacidad de absorción y retención de agua en el suelo (FAO 1991; Eduardo et al.
�����
���������� �����
�����
�����
����
��
��
��
��
������������
������
�� � � � � � �
�
Figura 1. Efecto de los tratamientos sobre la ca-pacidad de absorción de agua en el suelo.
����
������������
������
����
����
����
����
����
����
� � � � � � �
������
������������ ������
������
������
������
Figura 2. Efecto de los tratamientos sobre la densidad aparente del suelo.
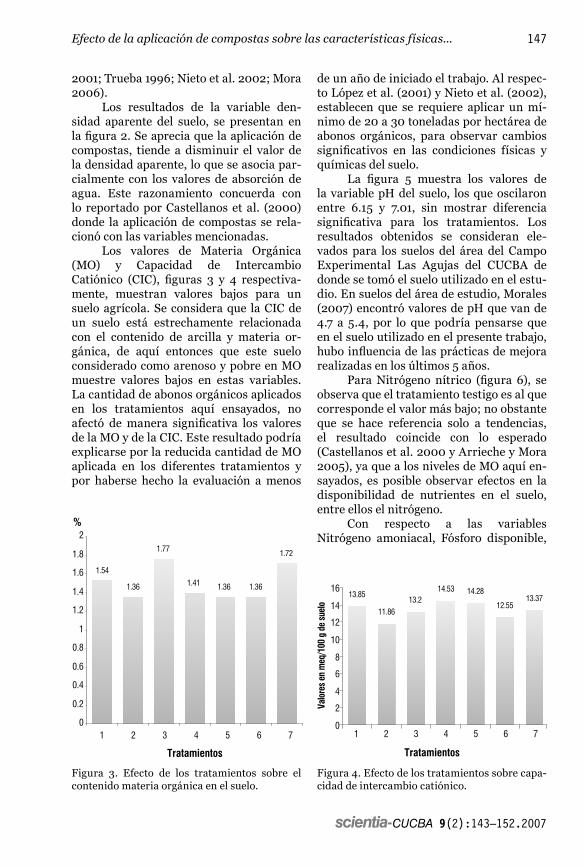
Efecto de la aplicación de compostas sobre las características físicas... 147
scientia-CUCBA 9(2):143—152.2007
2001; Trueba 1996; Nieto et al. 2002; Mora 2006).
Los resultados de la variable den-sidad aparente del suelo, se presentan en la figura 2. Se aprecia que la aplicación de compostas, tiende a disminuir el valor de la densidad aparente, lo que se asocia par-cialmente con los valores de absorción de agua. Este razonamiento concuerda con lo reportado por Castellanos et al. (2000) donde la aplicación de compostas se rela-cionó con las variables mencionadas.
Los valores de Materia Orgánica (MO) y Capacidad de Intercambio Catiónico (CIC), figuras 3 y 4 respectiva-mente, muestran valores bajos para un suelo agrícola. Se considera que la CIC de un suelo está estrechamente relacionada con el contenido de arcilla y materia or-gánica, de aquí entonces que este suelo considerado como arenoso y pobre en MO muestre valores bajos en estas variables. La cantidad de abonos orgánicos aplicados en los tratamientos aquí ensayados, no afectó de manera significativa los valores de la MO y de la CIC. Este resultado podría explicarse por la reducida cantidad de MO aplicada en los diferentes tratamientos y por haberse hecho la evaluación a menos
����
����
����
���� ���� ����
����
������������
�
� � � � � � �
�
�
���
���
���
���
�
���
���
���
���
Figura 3. Efecto de los tratamientos sobre el contenido materia orgánica en el suelo.
�����
���������
����� �����
����������
������������
�����������������������������
��
�
�
�
�
�
��
��
��
� � � � � � �
Figura 4. Efecto de los tratamientos sobre capa-cidad de intercambio catiónico.
de un año de iniciado el trabajo. Al respec-to López et al. (2001) y Nieto et al. (2002), establecen que se requiere aplicar un mí-nimo de 20 a 30 toneladas por hectárea de abonos orgánicos, para observar cambios significativos en las condiciones físicas y químicas del suelo.
La figura 5 muestra los valores de la variable pH del suelo, los que oscilaron entre 6.15 y 7.01, sin mostrar diferencia significativa para los tratamientos. Los resultados obtenidos se consideran ele-vados para los suelos del área del Campo Experimental Las Agujas del CUCBA de donde se tomó el suelo utilizado en el estu-dio. En suelos del área de estudio, Morales (2007) encontró valores de pH que van de 4.7 a 5.4, por lo que podría pensarse que en el suelo utilizado en el presente trabajo, hubo influencia de las prácticas de mejora realizadas en los últimos 5 años.
Para Nitrógeno nítrico (figura 6), se observa que el tratamiento testigo es al que corresponde el valor más bajo; no obstante que se hace referencia solo a tendencias, el resultado coincide con lo esperado (Castellanos et al. 2000 y Arrieche y Mora 2005), ya que a los niveles de MO aquí en-sayados, es posible observar efectos en la disponibilidad de nutrientes en el suelo, entre ellos el nitrógeno.
Con respecto a las variables Nitrógeno amoniacal, Fósforo disponible,

148 Martha Delia Ocegueda Reyes et al.
scientia-CUCBA 9(2):143—152.2007
Potasio, relación Carbono Nitrógeno (C/N) y conductividad eléctrica (CE), los trata-mientos aplicados al suelo no mostraron diferencia significativa. Como se mencionó en párrafos anteriores, este resultado pue-de obedecer a la baja cantidad de composta aplicada, y a que los datos se tomaron antes de que se cumpliera un año de la incorpo-ración de los materiales, ya que se afirma que la aplicación de niveles altos de MO con un suficiente periodo de incubación
debe incrementar los valores correspon-dientes a estas variables (Castellanos et al. 2000).
Los resultados de los análisis de varianza referentes a las variables mi-crobiológicas de hongos, bacterias y ac-tinomicetos, reportadas como Unidades Formadores de Colonias (UFC), mostraron diferencias significativas para bacterias (p < 0.01) y actinomicetos (p < 0.05) (cua-dro 3).
����
����
����
��������
����
����
������������
��
� � � � � � ����
���
�
���
���
���
���
�
���
Figura 5. Efecto de los tratamientos sobre el pH del suelo.
��
��
��
�� �� ��
��
������������
���
� � � � � � ��
��
��
��
��
��
��
�
Figura 6. Efecto de los tratamientos sobre el contenido de N- NO3 en el suelo.
Cuadro 3. Prueba de separación de medias para unidades formadoras de colonias de actinomicetos y bacterias (UFC x 106).
No. de tratamiento
Tratamientos Actinomicetos promedio
(UFC x 106)
Actinomicetos Tukey
(p <0.05)
Bacterias promedio
(UFC x 106)
Bacterias Tukey (p <0.05)
1 Composta tipo
Bocashi
20.0 b 35 b
2 Composta paja
frijol
165.0 a 107.5 a
3 Composta paja
maíz + frijol
20.0 b 75 a
4 Composta maíz 70.0 a 52.5 b
5 Vermiabono 40.0 b 32.5 b
6 Fertilizante 42.5 b 40 b
7 Testigo 40.0 b 47.5 b
Cifras seguidas de la misma letra indican igualdad estadística.

Efecto de la aplicación de compostas sobre las características físicas... 149
scientia-CUCBA 9(2):143—152.2007
La población microbiana del suelo se estimula con la adición de materia orgáni-ca; al mantener la actividad microbiana, se activa el ciclo del carbono y con ello el buen funcionamiento del agroecosistema (López-Alcocer et al. 2000). De acuerdo con la literatura, la población de hongos en suelos agrícolas varía de 20,000 hasta 1’000,000 UFC, las bacterias de unos cuan-tos miles hasta 200 millones, en tanto que para actinomicetos han reportado valores que van desde 30 a 2380 × 103 (Alexander 1977; Valdés et al. 1980). Si se consideran las limitaciones de los métodos existentes para cuantificar la población microbiana en el suelo, se colige que los valores en-contrados en el presente estudio, se ubican dentro de los parámetros esperado para un suelo agrícola.
Respecto a la población de actinomi-cetos (cuadro 3), la prueba de separación de medias (Tukey, p <0.05) mostró que los tratamientos 2 (composta de frijol) y 4 (composta de maíz) fueron superiores al resto de tratamientos ensayados, en tan-to que para bacterias, los tratamientos 2 (composta de frijol) y 3 (composta de maíz-frijol) son iguales entre sí y superiores al resto de los tratamientos (cuadro 3). En el presente estudio, el desarrollo microbiano, al parecer, se ve favorecido por la presencia de composta desarrollada a partir de paja de frijol, material con alto contenido de nitrógeno y bajo contenido en lignina, con
una relación C/N, menor a 40 (Stoffella y Kahn 2005).
Con respecto a las variables medidas en planta, los análisis de varianza detec-taron diferencias altamente significativas entre tratamientos para nodulación y peso de grano, no así para las variables longitud de parte aérea, longitud de raíz, peso seco parte aérea, peso seco de raíz y número de vainas.
Para el número de nódulos por plan-ta, los resultados de la prueba de separa-ción de medias se presentan en el cuadro 4. Se aprecia una tendencia clara de mayor número de nódulos en los tratamientos que llevaron aplicación de composta; la tendencia observada es congruente con los informes de Gutiérrez et al. (2001), quienes al aplicar composta al suelo, observaron un incremento significativo en número de nódulos en Vigna unguiculata L. Por otra parte, la prueba estadística detectó dife-rencias significativas entre el tratamiento 4 (composta de maíz) que registró el mayor número de nódulos y el tratamiento 6 (con fertilizante químico), con la menor nodu-lación. Este resultado es congruente con la literatura, pues está bien documentada la inhibición de la nodulación y la fijación de nitrógeno en frijol, cuando se aplican ferti-lizantes nitrogenados al suelo (Dart 1977).
En el caso de número de vainas por planta, el análisis de varianza no detectó diferencias significativas para tratamien-
Cuadro 4. Resultados de la prueba de separación de medias para la variable nodulación.
No. de tratamiento Tratamientos Nódulos por planta (promedio)
Tukey(p < 0.05)
4 Composta paja maíz 40.04 a
3 Composta paja frijol-maíz 39.23 ab
5 Vermiabono 37.06 ab
2 Composta paja frijol 24.72 ab
7 Testigo 22.45 ab
1 Composta tipo Bocashi 22.02 ab
6 Fertilizante 15.42 b
Cifras seguidas de la misma letra indican igualdad estadística.

150 Martha Delia Ocegueda Reyes et al.
scientia-CUCBA 9(2):143—152.2007
tos; sin embargo, alcanzó un valor de 0.0814 de probabilidad de error. Por lo anterior y si toma en cuenta que el núme-ro de vainas por planta es un componente del rendimiento y muestra una correlación con el peso de grano, se generó la ecuación de regresión (figura 7), que muestra que el 77% del peso de grano está determinado por el número de vainas. Es decir, la rela-ción entre las variables número de vainas por planta y peso de grano, es muy estre-cha y confirma que el número de vainas es uno de los componentes del rendimiento de grano más importantes en frijol (White e Izquierdo 1989).
La prueba de separación de medias (Tukey p < 0.05) para peso de grano por planta, se muestra en el cuadro 5. La mayo-ría de los tratamientos a base de composta, presentaron rendimientos estadísticamen-te iguales al tratamiento fertilizado; este resultado refleja que los materiales orgá-nicos son una opción viable para eliminar, o por lo menos disminuir el uso del ferti-lizante químico en la producción de frijol (Morales 2007). Por otra parte, el compor-tamiento del tratamiento testigo de valor similar al tratamiento fertilizado, no tiene lógica aparente en este estudio, el resulta-do podría obedecer a la heterogeneidad del suelo o algún error de toma de datos.
Adicionalmente, la figura 8 muestra la relación encontrada entre el conteni-do de Magnesio del suelo y la longitud de parte aérea de la planta; se observa que este nutrimento influyó de manera posi-tiva sobre el crecimiento (r = 0.57*). De acuerdo con este resultado, el contenido de Magnesio determinó el 32% de la longitud de la parte aérea de la planta de frijol. Un efecto positivo del Magnesio, ya sea del nativo del suelo o el que se aplique como fertilizante para el desarrollo de los cul-tivos, se presenta en suelos que han sido repetidamente encalados con materiales carentes de Magnesio (Jones 2003).
Conclusiones
Con base a los resultados obtenidos y bajo las circunstancias que prevalecieron durante el desarrollo del experimento, se deduce que:
• La aplicación de compostas al suelo mostró efectos sobre las características físicas del mismo, con una tendencia a favorecer la capacidad de absorción de agua y a disminuir la densidad aparen-te.
• El volumen de aplicación de compostas, no mostró efectos sobre las variables químicas de suelo consideradas.
��������������������
�����������
�������
������
�� � �� �� �� �� ��
�
��
��
��
��
��
��
��
Figura 7. Relación entre número de vainas por planta y el peso de grano por planta.
��������������������
�����������
�����������������������������
���
� �� �� �� �� �� ���
��
��
��
��
���
���
Figura 8. Relación entre Magnesio y longitud de parte aérea de la planta de frijol.

Efecto de la aplicación de compostas sobre las características físicas... 151
scientia-CUCBA 9(2):143—152.2007
• La aplicación de compostas al suelo mostró efectos sobre la población mi-crobiana de actinomicetos y bacterias, no así sobre la población de hongos.
• La planta de frijol mostró respuesta a la aplicación de compostas al suelo, en las variables nodulación y rendimiento de grano.
• Para inducir un mayor efecto en las características físicas, químicas y bioló-gicas del suelo, se requiere de la aplica-ción periódica de mayores volúmenes de composta.
• La aplicación de abonos orgánicos pue-de disminuir o sustituir la aplicación de fertilizante químico en la producción de grano en el cultivo de frijol.
Bibliografía
Alexander, M. 1977. Introduction to soil mi-crobiology. John Wiley and Sons. USA. Pp. 438–455.
Arrieche, I. y O. Mora. 2005. Efecto de la aplicación de residuos orgánicos sobre el cultivo de maíz en suelos degradados del estado de Yaracuy, Venezuela. Biagro 17(3): 155–159.
Avilés, G. M. y J. Tello. 2001. El composta-do de los residuos orgánicos, su relación con las enfermedades de las plantas. In: Agroecología y desarrollo. Labrador, M. J. y M.A. Altieri. (Coordinadores). Mundi-Prensa. España. Pp. 185–216.
Castellanos, J. Z., X. Uvalle y A. Aguilar. 2000. Manual de interpretación de suelos y
aguas. Instituto de capacitación para la pro-ductividad agrícola. 2da. Edición. 226 pp.
Dalzell, H., W. J. Biddlestone, R. Gray y K. Thurairajan. 1991. Manejo del suelo: producción y uso del composta en ambien-tes tropicales y subtropicales. Servicio de recursos, manejo y conservación del suelo. Dirección de fomentos de tierras y aguas. FAO. Roma. 178 pp.
Dart, P. 1977. Infection and development of luguminous nodules. In: A treatise on di-nitrogen fixation, Section III. Hardy R.W.F. Editor.. John Wiley and Sons. USA. Pp. 367–472.
Eduardo, A., L. Tijerina, A. Acosta y A. López. 2001. Producción de ciruelo con fertirriego en función de contenidos de hu-medad y coberturas orgánicas. Terra 19: 317–326.
FAO. 1991. Manejo del suelo producción y uso de composta en ambientes tropica-les. Boletín de Suelos. Organización de las Naciones Unidas para la Agricultura y la Alimentación. 178 pp.
Fisher, P., E. Miramontes y M. Crespo. 2003. Manual de practicas del curso “Uso y manejo de compostas para sustratos de hortalizas y ornamentales”. Centro Universitario de Ciencias Biológicas y Agropecuarias, Universidad de Guadalajara, México. Pp. 64–71.
Gutiérrez, W., C. Medrano, M. Materan, Y. Villalobos, D. Esparza, J. Baéz y B. Medina. 2001. Evaluación del rendimiento y nodulación del frijol Vigna unguiculata (L.) Walp bajo dos sistemas de labranza en las condiciones agroecológicas de la planicie de Maracaibo, Venezuela. Rev. Fac. Agron. 18: 237–246.
Cuadro 5. Resultados de la prueba de separación de medias para la variable peso de grano.
No. de tratamiento Tratamientos Promedio (g por planta) Tukey (p < 0.05)
5 Vermiabono 23.3820 a
7 Testigo 22.3880 ab
6 Fertilizante 17.4980 ab
3 Composta paja frijol-Maíz 16.2700 ab
2 Composta paja frijol 14.4120 ab
4 Composta paja maíz 13.9480 ab
1 Composta tipo Bocashi 6.4820 b
Cifras seguidas de la misma letra indican igualdad estadística.

152 Martha Delia Ocegueda Reyes et al.
scientia-CUCBA 9(2):143—152.2007
Havlin, J. L., J. D. Beaton, S. L. Tisdale and W. L. Nelson. 2005. Soil fertility and fertilizers. An introduction to nutrient ma-nagement. Seventh Edition. Prentice Hall, Upper Saddle River, NJ, USA. 515 pp.
Jones, J. and J. Benton. 2003. Management of crops, soils, and their fertility. Agronomic Handbook. CRC Press. 450 p.
Kononova, M. 1965. Soil Organic Matter Its nature, its role in soil formation and in soil fertility. Pergamon Press 2ad. Edition.
Labrador, M. J. 1996. La materia Orgánica en los agroecosistemas. Mundi-Prensa. España. 174 pp.
López, M., A. Díaz, E. Martínez y D. Valdez. 2001. Abonos orgánicos y su efecto en propiedades físicas y químicas del suelo y rendimientos de maíz. Terra 19: 293–299.
Lopez-Alcocer, E., P. Topete, F. Calderón, C. y R. Canales. 2000. El sistema de pro-ducción: un elemento para la generación de tecnología en la disciplina agroecológica. Scientia-CUCBA 2: 48–59.
López, A. E. y F. López. 2004. Ecosistemas y Agroecosistemas. In: Agroecología: Principios y Métodos. E. López-Alcocer, S. Hurtado y F. López-Alcocer (Editores). Universidad de Guadalajara. México. Pp. 7–12.
Luna de, V. A. y E. Vázquez. 2003. Elaboración de abonos orgánicos. Universidad de Guadalajara. México.
Mora, D. J. 2006. Contribuciones del compost al mejoramiento de la fertilidad del suelo. Universidad de Costa Rica. http://lunazul.ucaldas.edu.co
Morales T., M. 2007. Efecto de la aplicación de agroquímicos y abonos orgánicos so-bre la producción de frijol y la población microbiana del suelo. Tesis de maestría en Ciencias Agrícolas y Forestales. CUCBA. Universidad de Guadalajara. 80 pp.
Nieto, G. A., E. Troyo, B. Murillo, J. García y A. Larrinaga. 2002. La com-posta. Importancia, elaboración y uso agrí-cola. Centro de investigaciones biológicas del Noroeste. S. C. La Paz, BCS. México. Pp. 6–57.
Queitsch, K. J. 2005. Necesidad de desarro-llar la agricultura ecológica. In: Agricultura ecológica y desarrollo regional sustenta-ble. Producir en armonía con la naturaleza. Promotores de la autogestión para el desa-
rrollo social S. C. Universidad del Sur A. C. México. 448 pp.
Richards, L. A. 1982. Diagnóstico y rehabili-tación de suelos salinos y sódicos. Limusa. México. Cuarta reimpresión. 172 pp.
Steel, G. D. y J. Torrie. 1985. Bioestadísticas. Principios y procedimientos. Mc-Graw-Hill. Latinoamericana, S. A. 2da. Edición. Colombia. 622 pp.
Stoffella, P. J. y A. Kahn. 2005. Utilización de compost en los sistemas de cultivo hortí-cola. Mundi-Prensa. México. 397 pp.
Tcobanoglous, G., H. Theisen y S. Vigil. 1994. Gestión integral de residuos sólidos. Vol. II. Mc Graw Hill. México. Pp. 775-806.
Torres, A., C. Rivero, Y. Ampueda y E. Cori de. 2002. Efecto de la incorporación de residuos orgánicos marcados con 15N so-bre la dinámica del nitrógeno en dos suelos venezolanos. Rev. Fac. Agron. (Maracay) 28: 105–116.
Triano, S. A., D. Palma y E. Hernández. 2005. Uso de sustratos orgánicos y reci-claje de nutrimentos en la producción de cacao (Theobroma cacao L.) en el municipio de Cunduacán, Tabasco. Colegio de post-graduados en ciencias agrícolas, campus Tabasco. Fundación PRODUCE, Tabasco, A.C.
Trueba, C., S. 1996. Fertilizantes Orgánicos y Compostas. In: Agricultura orgánica: Una opción sustentable para el agro mexicano. Universidad Autónoma Chapingo. México. 163 pp.
Valdés, R. M., A. Echegaray, V. Olalde, A. M. Vazquez, J. Rodríguez y J.J. Ortega. 2003. Manual practico de Microbiología agrícola. Instituto Politécnico Nacional, Escuela Nacional de Ciencias Biológicas. Departamento de Microbiología. 69 pp.
Velásquez, G. J. 1997. Avances de la investiga-ción en labranza de conservación. INIFAP. México. Pp. 3–10.
White, J. y J. Izquierdo. 1989. FRIJOL: Fisiología del potencial de rendimiento y la tolerancia al estrés. CIAT-FAO. Santiago, Chile. 91 pp.
FECHA EFECTIVA DE PUBLICACIÓN 29 DE DICIEMBRE DE 2007


















